生物学 第2版 — 第22章 原核生物:細菌と古細菌 —
Japanese translation of “Biology 2e”
OpenStax のサイトで公開されている教科書“ Biology 2e”の翻訳です。こちらのページから各章へ移動できます。
22 | 原核生物:細菌と古細菌

この章の概要
22.1:原核生物の多様性
22.2:原核生物の構造:細菌と古細菌
22.3:原核生物の代謝
22.4:人間における細菌性疾患
22.5:有益な原核生物
はじめに
つい最近まで、科学者たちは、核や他の膜に包まれた細胞小器官の存在・不存在、細胞壁の存在・不存在、多細胞性など、いくつかの基準に基づいて生物を5つの界 — 動物、植物、菌類、原生生物、原核生物 — に分類していました。20世紀後半、カール・ウーズと他の者たちの先駆的な研究は、小サブユニットリボソームRNA(SSU rRNA)の配列を比較し、それは地球上の生物をグループ化するためのより基本的な方法をもたらしました。細胞膜の構造とrRNAの違いに基づいて、ウーズと彼の同僚たちは、地球上のすべての生命がドメインと呼ばれる3つの系統に沿って進化したことを提案しました。細菌のドメインは細菌界のすべての生物からなり、古細菌のドメインは原核生物の残りの部分からなり、真核生物のドメインは動物界、植物界、菌界、および原生生物界の生物を含むすべての真核生物からなります。
3つのドメインのうち2つ — 細菌と古細菌 — は原核生物です。原核生物は地球上で最初の住人で、35~38億年前に出現しました。これらの生物は豊富で遍在しています。つまり、彼らはいたるところに存在しています。穏やかな環境に生息することに加えて、それらは極端な条件でも発見されています:沸騰する泉から南極の永久に凍った環境まで、死海のような塩分の多い環境から深海のような途方もない圧力のかかる環境まで、そして廃棄物処理プラントなどの酸素のない領域からチェルノブイリなどの放射能で汚染された地域まで。原核生物は人間の消化器系や皮膚に存在したり、特定の病気の原因となったり、多くの食品の調理において重要な役割を果たしたりしています。
22.1 | 原核生物の多様性
この節が終わるまでに、あなたは次のことができるようになります:
•原核生物の進化の歴史を記述する
•好極限性生物の際立った特徴について議論する
•原核生物を培養するのが難しい理由を説明する
原核生物はいたるところにいます。それらは十分な湿気のある考えられるすべての表面を覆っています。そしてそれらはまた、実質的に他のすべての生物の表面上とその内部にも住んでいます。典型的な人体において、原核細胞は人間の体細胞よりも約10倍多いです。それらはすべての生態系の生物の大部分を構成しています。原核生物の中には、ほとんどの生物にとって生存に適さない環境で繁栄するものがあります。原核生物は栄養素 — 必須の物質(炭素や窒素など) — をリサイクルし、それらは新しい生態系(そのうちのいくつかは自然のもので、他のものは人工のものです)の進化を推進します。原核生物は、多細胞生物が出現するずっと前から地球上に存在しています。実際のところ、真核細胞は古代の原核生物の生物群集の子孫であると考えられています。
原核生物、地球の最初の住民
いつ、どこで細胞生物が始まったのでしょうか?生命が始まったときの地球の状況はどうだったのでしょうか?私たちは、原核生物がおそらく地球上での細胞生物の最初の形であり、植物や動物が出現するまでには何十億年もの間存在していた、ということを知っています。地球と月の年齢は約45.4億年です。この推定値は、隕石物質と地球および月からの他の基質物質の放射性年代測定からの証拠に基づいています。初期の地球は、今日とは非常に異なる大気(含まれる酸素分子はより少ない)を持っていて、強い太陽放射を受けていました。したがって、最初の生物はおそらく、深海や地球の表面のはるか下など、より保護された場所で繁栄していたでしょう。この時点では地球上では強い火山活動は一般的でした。従ってこれらの最初の生物 — 最初の原核生物 — は非常な高温に適応した可能性が高いです。初期の地球は地質学的激動と火山噴火を起こしやすく、太陽からの変異原性放射線にさらされていたので、最初の生物はこれらの過酷な条件に耐えなければならないような原核生物でした。
微生物マット
微生物マットまたは大きなバイオフィルムは、地球上で最も初期の原核生物の形態を表しているかもしれません。およそ35億年前に始まったそれらの存在の化石証拠があります。地球自体が形成されてからわずか10億年後に細胞生物が地球に現れたことは注目に値します。それは、自身を複製することができる細胞前の「生命」がはるかに早く進化したことを示唆しています。微生物マットは、ほとんどが細菌ですが古細菌も含むような原核生物の多層シートです(図22.2)。微生物マットは厚さがわずか数センチメートルであり、それらは典型的には異なる種類の材料が接触する場所(ほとんどが湿った表面上)で成長します。それらを構成するさまざまな種類の原核生物は異なる代謝経路を実行します。それが、微生物マットのさまざまな色の理由です。微生物マット内の原核生物は、それらが分泌する細胞外基質と呼ばれる接着剤のような粘着性物質によって一緒に保持されています。
最初の微生物マットは、熱水噴出孔の近くにある化学物質からエネルギーを得ていたようです。熱水噴出孔は、地表面の割れ目または亀裂であり、地熱によって加熱された水を放出します。約30億年前の光合成の進化に伴い、微生物マットの中の原核生物の中にはより広く利用可能なエネルギー源である太陽光を使用するようになったものもありました。一方で他のものは、エネルギーと食事をまだ熱水噴出孔からの化学物質に依存していました。

ストロマトライト
化石化した微生物マットは、地球上で最も初期の生命の記録を表しています。ストロマトライトは、微生物マット内の原核生物によってミネラルが水から析出するときに形成される堆積構造です(図22.3)。ストロマトライトは炭酸塩またはケイ酸塩からなる層状の岩を形成します。ほとんどのストロマトライトは過去の遺物ですが、ストロマトライトがまだ形成されている場所が地球上にはいくつかあります。たとえば、カリフォルニア州サンディエゴ郡のアンザ-ボレゴ砂漠州立公園で、成長しているストロマトライトが発見されました。

古代の大気
証拠は、地球が存在した最初の20億年の間は大気が無酸素、つまり分子状の酸素が存在しなかったことを示しています。したがって、酸素なしで成長することができる生物、嫌気性生物だけが生きることができました。太陽エネルギーを化学エネルギーに変換する独立栄養生物は光栄養生物と呼ばれ、それらは地球の形成から10億年以内に出現しました。そして、「藍藻」としても知られるシアノバクテリアは、少なくとも10億年後にこれらの単純な光栄養生物から進化しました。大気の「酸素化」を開始したのは、祖先のシアノバクテリア(図22.4)でした。大気中の酸素が増加すると、より効率的なO₂を利用した異化経路の進化が可能になりました。またそれはさらなる生息地としての地上への道を開きました。なぜなら、一部のO₂はO₃(オゾン)に変換され、オゾンはDNAに致命的な突然変異を引き起こす可能性がある紫外線を効果的に吸収するからです。現在の証拠は、O₂濃度の増加が他の形態の生物の進化を可能にしたことを示唆しています。

微生物は適応可能である:穏やかな環境と極端な環境での生命
いくつかの生物は、過酷な条件を乗り切ることを可能にする戦略を発展させました。ほとんどすべての原核生物は細胞壁、すなわち高張性および低張性の両方の水性条件で生き残ることを可能にする保護構造を有します。いくつかの土壌細菌は、熱や干ばつに抵抗する内生胞子を形成することができ、それによって好ましい条件が再び起きるまでこの生物が生き残ることを可能にします。これらの適応は、他のものと一緒になって、すべての陸上生態系および水中生態系において細菌が最もありふれた生命の形態であり続けることを可能にします。
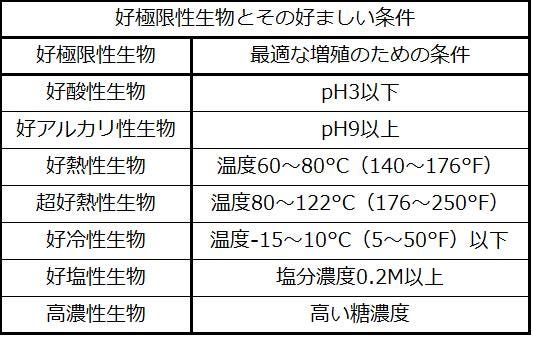
原核生物は幅広い環境の中で繁栄します。あるものは私たちにとって非常に普通に見える条件で育ちますが、他のものは植物や動物を殺すような条件のもとで繁栄し育つことができます。極限環境下で増殖するように適応した細菌と古細菌は、「極限環境を愛するもの」を意味する好極限性生物と呼ばれています。好極限性生物はあらゆる種類の環境で発見されています:ほんの数例を挙げると、深海、温泉、北極と南極、極寒、極度に乾燥した場所、地球の奥深く、過酷な化学的環境、および高放射線環境です(図22.5)。それらは極端な条件で生きることを可能にする特別な適応を持っているので、多くの好極限性生物は穏やかな環境では生き残ることができません。好極限性生物にはさまざまなグループがあります。それらは、最もよく成長する条件に基づいて識別され、いくつかの生息地は複数の点で極端なものです。たとえば、ソーダ湖は塩分とアルカリ性の両方であるため、ソーダ湖に生息する生物は好アルカリ性と好塩性の両方でなければなりません(表22.1)。放射線耐性生物のような他の好極限性生物は極端な環境(この場合、高レベルの放射線を伴う環境)を好むわけではありませんが、その中で生き残るために適応してきました(図22.5)。このような生物は私たちに原核生物の多様性についてのより良い理解を与え、そして新しい治療薬の発見につながるか、または産業上の応用があるかもしれない新しい原核生物の種を見つける可能性を開きます。
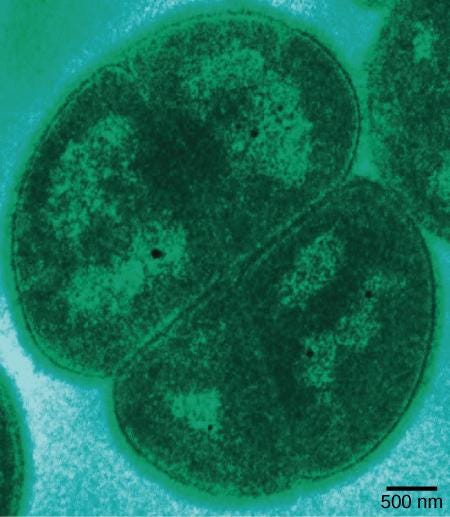
死海の原核生物
非常に過酷な環境の一例は、ヨルダンとイスラエルの間に位置する高塩分の湖である死海です。高塩分環境とは、本質的には濃縮された海水です。死海では、ナトリウム濃度は海水よりも10倍高く、水には高レベルのマグネシウムが含まれています(海水よりも約40倍高いです)。これは、ほとんどの生物にとって有害です。二価イオン(Fe²⁺、Ca²⁺、およびMg²⁺)を形成する元素である鉄、カルシウム、およびマグネシウムは、一般に「硬」水と呼ばれるものを生成します。まとめると、高濃度の2価のカチオン、酸性のpH(6.0)、そして強い太陽放射束によって、死海は独特かつ他に類を見ないほど厳しい生態系になります[1](図22.6)。
[1] Bodaker, I, Itai, S, Suzuki, MT, Feingersch, R, Rosenberg, M, Maguire, ME, Shimshon, B, and others. Comparative community genomics in the Dead Sea: An increasingly extreme environment. The ISME Journal 4 (2010): 399–407, doi:10.1038/ismej.2009.141. published online 24 December 2009.
私たちは、死海にどのような原核生物を見つけることができるでしょうか?極端に耐塩性に優れた細菌マットには、とりわけ、好塩菌のハロフェラックス・ヴォルカニイ(Haloferax volcanii)(死海だけでなく他の場所にもいます)、ハロルブラム・ソドメンス(Halorubrum sodomense)、ハロバクラム・ゴモレンス(Halobaculum gomorrense)、そして古細菌のハロアークラ・マリスモルティ(Haloarcula marismortui)があります。
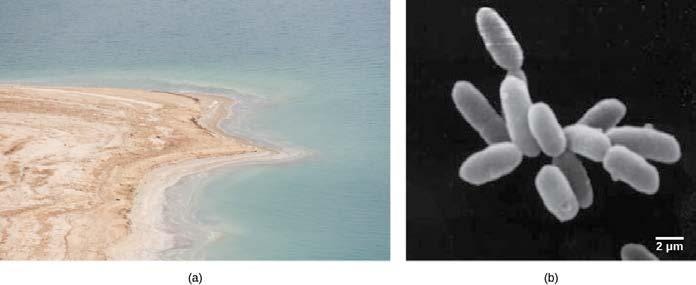
培養不可能な原核生物と生存可能だが培養不可能な状態
細菌を培養するプロセスは複雑であり、現代科学の最大の発見の1つです。ドイツの医師ロベルト・コッホは、染色や増殖培地の使用などを含む純粋培養のための技術を発見したと認められています。微生物学者は通常、標的となる生物に必要なすべての栄養素を含む適切な培地を用いて実験室で原核生物を増殖させます。培地は液体、煮出し汁、または固体であり得ます。適切な温度での培養時間の後には、微生物の増殖の証拠があるはずです(図22.7)。コッホの助手ユリウス・ペトリは、ペトリ皿を発明しました。ペトリ皿の使用は今日の研究室でも続いています。コッホは、結核を引き起こす結核菌(Mycobacterium tuberculosis)を用いて主に作業し、特定の病気の原因となる生物を識別するためのコッホの原則と呼ばれる指針を作成しました。コッホの原則は医学界で広く使われ続けています。コッホの原則は、ある生物が感染したすべてのサンプルに存在する、すべての健康なサンプルには存在しない、および複数回培養した後に感染を再現できるときに、その生物が疾患の原因として特定できるということを含むものです。今日、培養は医学や分子生物学のその他の分野における主要な診断ツールであり続けています。
コッホの原則は、単離および培養が可能な生物にのみ完全に適用することができます。しかしながら、原核生物の中には実験室の環境では成長できないものがあります。事実、細菌と古細菌の99%以上が培養不可能です。これはほとんどの場合、これらの生物に何を与えるべきか、そしてそれらをどのように成長させるかに関する知識の欠如によるものです。それらは、特定の微量栄養素、pH、温度、圧力、補助因子、または共代謝物を必要とするなど、科学者には知られていない、成長のための特別な要件があるのかもしれません。いくつかの細菌は、偏性細胞内寄生生物であり、宿主細胞の外で増殖することができないために培養することができません。
他の場合では、培養可能な生物は、たとえその同じ生物を以前に培養することができていたとしても、ストレスの多い条件下で培養不可能になります。培養することができないものの死んでいない生物は、生存可能だが培養不可能(VBNC)状態にあります。原核生物が環境ストレス要因に対して、その生物を生存可能にする休眠状態に入ることによって反応するとき、VBNC状態が起こります。VBNC状態に入るための基準は完全には理解されていません。蘇生と呼ばれるプロセスでは、原核生物は環境条件が改善されたときに「通常の」生活に戻ることができます。
VBNC状態は原核生物にとって珍しい生き方なのでしょうか?実際のところ、土壌や海水に生息する原核生物のほとんどは培養不可能です。実験室条件下では、原核生物のほんのわずかな割合、おそらく1%しか培養することができないと言われてきました。もしこれらの生物が培養不可能であるならば、それらが存在しそして生きているかどうかはどうやって知ることができるのでしょうか?微生物学者は、ポリメラーゼ連鎖反応(PCR)などの分子技術を使用して原核生物のDNAの選択された部分、たとえば16S rRNA遺伝子を増幅し、それらの存在を実証します。(PCRは増幅と呼ばれるプロセスでDNAセグメントの何十億ものコピーを作ることができることを思い出してください。)
バイオフィルムの生態
いくつかの原核生物は、他の原核生物種の存在を必要とするために培養不可能なのかもしれません。数十年前までは、微生物学者は原核生物のことを離れて住む孤立した存在として考えていました。しかしながら、このモデルは原核生物の真の生態を反映していません。原核生物のほとんどはそれらが相互作用することができる生物群集に住むことを好みます。私たちがこれまで見てきたように、バイオフィルムは、主に生物から分泌された多糖類といくつかのタンパク質や核酸からなる、ゴム状の質感のある基質にまとめられた微生物群集です(図22.8)。バイオフィルムは通常、表面に付着して成長します。最もよく研究されているバイオフィルムのいくつかは原核生物で構成されていますが、菌類バイオフィルムや菌類と細菌の混合物で構成されているものも記述されています。
バイオフィルムはほとんどどこにでも存在します。それらはパイプの目詰まりを引き起こしたり、工業的な環境で表面に容易にコロニーを形成したりすることがあります。最近では、食品の細菌汚染の大規模な発生において、バイオフィルムが大きな役割を果たしてきました。またそれらは、キッチンカウンター、まな板、流し台、トイレなどの家庭環境の表面や、私たちの歯の表面などの人体上の場所にもコロニーを形成します。
バイオフィルムに住む生物間の相互作用は、その保護的な菌体外多糖(EPS)環境とあいまって、これらの生物群集を自由生活性(またはプランクトン様)の原核生物よりも強固にします。細菌を一緒に保持する粘着性物質は、ほとんどの抗生物質や消毒剤も排除し、バイオフィルム細菌をそれらのプランクトン様の細菌よりも強固にします。全体として、バイオフィルムは多くの一般的な殺菌の形態に耐性があるため、破壊するのが非常に困難です。
ビジュアルコネクション

自由浮遊性の細菌と比較して、バイオフィルム内の細菌はしばしば抗生物質および洗剤に対する耐性の増加を示します。あなたはなぜそうなのだと思いますか?
22.2 | 原核生物の構造:細菌と古細菌
この節が終わるまでに、あなたは次のことができるようになります:
•典型的な原核生物の基本構造を記述する
•古細菌と細菌の構造の重要な違いを記述する
原核細胞と真核細胞の間には多くの違いがあります。「原核生物」という名称は、原核生物が排除によって定義されることを示唆しています — それらは真核生物(すなわち細胞が核および他の内部の膜に包まれた細胞小器官を含む生物)ではないものです。しかしながら、すべての細胞は4つの共通の構造を持っています:原形質膜(細胞のバリアとして機能し、細胞をその環境から分離します)、細胞質(細胞内の有機分子と塩の複合溶液)、二本鎖DNAゲノム(細胞の情報の保管庫)、リボソーム(タンパク質合成が行われる場所)です。原核生物はさまざまな形をしていますが、多くは3つのカテゴリーに分類されます:球菌(球状)、桿菌(棒状)、およびらせん菌(らせん状)です(図22.9)。

原核細胞
原核生物は、膜に包まれた細胞小器官または他の内部の膜に包まれた構造を欠く単細胞生物であることを思い出してください(図22.10)。それらの染色体(通常は1つ)は、核様体と呼ばれる細胞の領域に位置する環状の二本鎖DNAの断片からなります。ほとんどの原核生物は、原形質膜の外側に細胞壁を持っています。細胞壁は保護層として機能し、それはこの生物の形状に関与しています。いくつかの細菌の種は細胞壁の外側に莢膜を有します。莢膜は、この生物が表面に付着するのを可能にし、脱水および食細胞による攻撃から保護し、そして病原体を私たちの免疫反応に対してより耐性にします。いくつかの種はまた、移動に使用される鞭毛や、他の細胞の表面を含むさまざまな表面への付着に使用される性繊毛を有します。染色体外DNAからなるプラスミドもまた、多くの種類の細菌や古細菌に存在します。

原核生物は2つの異なるドメイン、細菌と古細菌に分けられていることを思い出してください。これらは真核生物とともに生命の3つのドメインを構成しています(図22.11)。
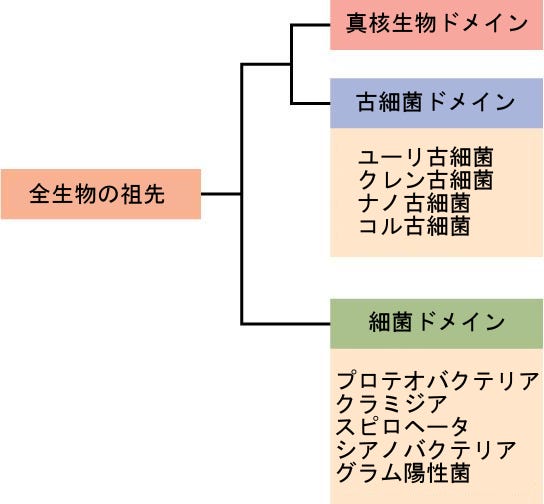
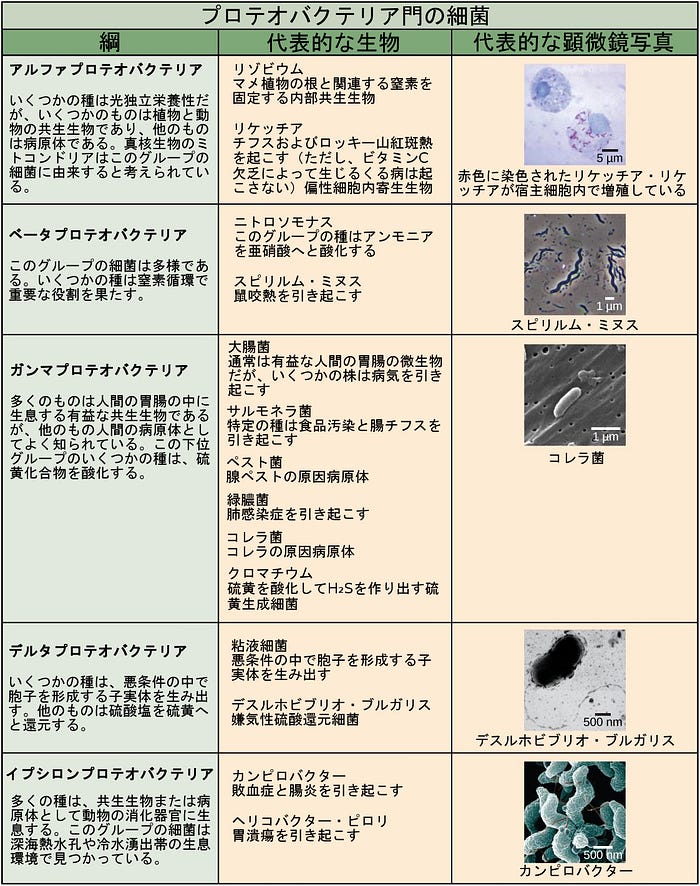
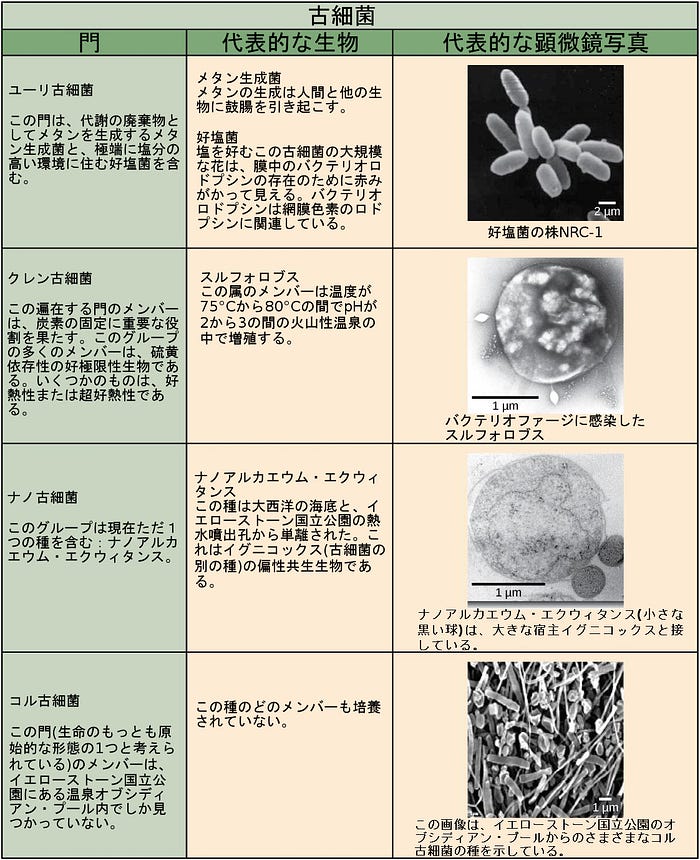
細菌の門の特徴が、図22.12および図22.13に記載されています。主な細菌の門は、プロテオバクテリア、クラミジア、スピロヘータ、光合成をするシアノバクテリア、およびグラム陽性菌を含みます。プロテオバクテリアは、次に、アルファプロテオバクテリアからイプシロンプロテオバクテリアまで、いくつかの綱に細分されます。真核生物のミトコンドリアは、アルファプロテオバクテリアの子孫である一方で、真核生物の葉緑体はシアノバクテリアに由来すると考えられています。古細菌の門は図22.14に記載されています。

原核生物の細胞膜
原核生物の原形質膜は、細胞を完全に囲み、内側と外側を隔てる薄い脂質二重層(6~8ナノメートル)です。その選択的透過性は、イオン、タンパク質、および他の分子を細胞内に保ち、それらが細胞外環境に拡散するのを防ぐ一方で、他の分子は膜を通って移動することができます。細胞膜の一般的な構造は、2つの層の脂質分子からなるリン脂質二重層であることを思い出してください。古細菌の細胞膜では、グリセロールに結合したイソプレン(フィタニル)鎖が、細菌の膜のグリセロールに結合した脂肪酸を置き換えています。いくつかの古細菌の膜は、二重層ではなく脂質単層です(図22.15)。

原核生物の細胞壁
原核細胞の細胞質は高濃度の溶解した溶質を有します。したがって、細胞内の浸透圧は比較的高いです。細胞壁とは、いくつかの細胞を囲み、それらに形状と剛性を与える保護層です。それは細胞膜の外側に位置し、浸透圧溶解(体積の増加による破裂)を防止します。細胞壁の化学組成は古細菌と細菌の間で異なり、また細菌の種の間でも異なります。
細菌の細胞壁は、ペプチドグリカン(L-およびD-アミノ酸(D-グルタミン酸およびD-アラニンを含む)の両方を含む非天然ペプチドによって架橋されている多糖鎖から構成される)を含みます。(タンパク質は通常L-アミノ酸しか有しません。その結果、私たちの抗生物質の多くはD-アミノ酸を模倣することによって働き、それゆえ細菌の細胞壁発達に特異的な効果をもたらします。)100以上の異なる形態のペプチドグリカンがあります。また、古細菌と細菌の両方の細胞壁の外側には、S層(表面層)タンパク質も存在します。
細菌は、グラム染色に対する反応に基づいて、グラム陽性とグラム陰性の2つの主要なグループに分けられます。グラム陽性菌はすべて1つの門に属します。他の門(プロテオバクテリア、クラミジア、スピロヘータ、シアノバクテリアなど)の細菌はグラム陰性です。グラム染色法は、その発明者であるデンマークの科学者ハンス・クリスチャン・グラム(1853–1938)にちなんで名付けられました。この染色手法に対する異なる細菌反応は、究極的には細胞壁構造によるものです。グラム陽性の生物には通常、グラム陰性の生物に見られる外膜がありません(図22.16)。グラム陽性菌の細胞壁の最大90%がペプチドグリカンで構成され、残りのほとんどがテイコ酸と呼ばれる酸性物質で構成されています。テイコ酸は、原形質膜中の脂質と共有結合してリポテイコ酸を形成することがあります。リポテイコ酸は細胞壁を細胞膜に固定します。グラム陰性菌は、リポ多糖類(LPS)およびリポタンパク質を含む外側エンベロープに囲まれた、数層のペプチドグリカン(全細胞壁のわずか10%)からなる比較的薄い細胞壁を有します。この外側エンベロープは時に第二脂質二重層と呼ばれます。しかしながら、この外側エンベロープの化学的性質は、原形質膜を形成する典型的な脂質二重層の性質とは非常に異なります。
ビジュアルコネクション

次の記述のうち、正しいものはどれですか?
a.グラム陽性菌は、リポテイコ酸によって細胞膜に固定された単一の細胞壁を有する。
b.ポリンはグラム陽性菌とグラム陰性菌の両方への物質の侵入を可能にする。
c.グラム陰性菌の細胞壁は厚く、グラム陽性菌の細胞壁は薄い。
d.グラム陰性菌はペプチドグリカンからなる細胞壁を有するのに対し、グラム陽性菌はリポテイコ酸からなる細胞壁を有する。
古細菌の細胞壁はペプチドグリカンを持っていません。古細菌の細胞壁には4つの異なる種類があります。1つの種類は、疑似ペプチドグリカン(形態はペプチドグリカンに類似しているものの、多糖鎖中に異なる糖を含みます)から構成されます。他の3つの種類の細胞壁は、多糖類、糖タンパク質、または純粋なタンパク質で構成されています。細菌と古細菌の他の違いは表22.2に示されています。古細菌におけるDNA複製、転写および翻訳に関連する特徴は、真核生物において見られるものと類似していることに注意してください。

生殖
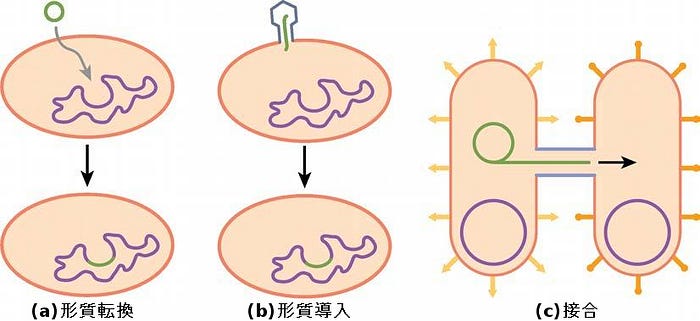
原核生物における生殖は無性生殖であり、通常二分裂によって行われます。(原核生物のDNAは単一の環状染色体であることを思い出してください。)原核生物は有糸分裂を起こしません。その代わりに、染色体が複製され、そして細胞の成長のために得られた2つのコピーが互いに分離します。その時点で拡大した原核生物は、赤道で内側にくびれて、クローンである2つの細胞が分離します。二分裂は遺伝子組換えや遺伝的多様性の機会を提供しませんが、原核生物は他の3つのメカニズムによって遺伝子を共有することができます。
形質転換において、原核生物は他の原核生物によって環境へとこぼされたDNAを取り込みます。もし非病原性細菌が病原体から毒素遺伝子のDNAを取り込み、それ自身の染色体に新しいDNAを取り込むと、それもまた病原性になるかもしれません。形質導入において、バクテリオファージ(細菌に感染するウイルス)は、染色体DNAの短い破片をある細菌から別の細菌に移動させることがあります。形質導入は組換え生物をもたらします。古細菌には、遺伝物質をある個体から別の個体に移動させることのあるウイルスもあります。接合において、DNAは、性繊毛(生物を互いに接触させ、そしてDNAの移動のためのチャネルを提供します)によってある原核生物から別の原核生物に移動されます。移入されるDNAは、プラスミドの形態でも、プラスミドと染色体DNAの両方を含む複合分子としてであってもかまいません。これら3つのDNA交換プロセスが図22.17に示されています。
生殖は非常に速いことがあります:種によっては数分です。この短い世代間の時間は、遺伝的組換えと高い率の突然変異のメカニズムと組み合わされて、原核生物の急速な進化をもたらし、それらが環境変化(抗生物質の導入など)に非常に迅速に反応することを可能にします。
進化へのつながり
原核生物の進化
科学者は原核生物の進化についての質問にどのように答えるのでしょうか?動物とは異なり、原核生物の化石記録にある遺物はほとんど情報を提供しません。古代の原核生物の化石は岩の中の小さな泡のように見えます。科学者の中には遺伝学と分子時計の原理(2つの種が分かれるのが最近であればあるほど、それらの遺伝子(したがってタンパク質)はより類似していると考えるものです。)に目を向けている人もいます。逆に、ずっと前に分岐した種は、似ていない遺伝子をより多く持つようになるでしょう。
NASA宇宙生物学研究所と欧州分子生物学研究所の科学者らは、72種の原核生物に共通する32個の特定のタンパク質の分子進化を分析するために共同研究を行いました[2]。彼らがデータから導いたモデルは、細菌の3つの重要なグループ — 放線菌、デイノコッカス、およびシアノバクテリア(著者たちによってまとめてテッラバクテリアと呼ばれています) — が最初に陸地に移住したことを示しています。放線菌は、菌類の菌糸のような分岐構造を作り出す、非常に一般的なグラム陽性菌のグループで、有機廃棄物の分解にとって重要な種を含みます。あなたは、デイノコッカスが電離放射線に非常に強い耐性を持つ細菌の属であることを覚えているでしょう。それは第2の外膜に加えて厚いペプチドグリカン層を有するので、グラム陽性菌およびグラム陰性菌の両方の特徴を有しています。
シアノバクテリアは光合成を行うものであり、おそらく古代の地球上での酸素の生産を担っていました。分岐の時系列は、細菌(細菌ドメインのメンバー)が25~32億年前に共通の祖先の種から分岐したのに対し、古細菌はそれより早く(31~41億年前)に分岐したことを示唆しています。真核生物は後に古細菌の線から分岐しました。この研究はさらに、シアノバクテリアの出現(約26億年前)よりも前に形成されたストロマトライトが無酸素環境で光合成していたこと、そして陸地に上がるためのテッラバクテリアの改変(乾燥に対する耐性およびこの生物を過剰な光から保護するための化合物の保有)の理由から、酸素を使った光合成は陸上で生き残るための適応に密接に関連しているかもしれないこと、を示唆しています。
[2] Battistuzzi, FU, Feijao, A, and Hedges, SB. A genomic timescale of prokaryote evolution: Insights into the origin of methanogenesis, phototrophy, and the colonization of land. BioMed Central: Evolutionary Biology 4 (2004): 44, doi:10.1186/1471–2148–4–44.
22.3 | 原核生物の代謝
この節が終わるまでに、あなたは次のことができるようになります:
•原核生物が必要とする主要栄養素を特定し、その重要性を説明する
•原核生物が生命プロセスのためにエネルギーと炭素を得る方法を記述する
•炭素と窒素の循環における原核生物の役割を記述する
原核生物は代謝的に多様な生物です。多くの場合、原核生物はその代謝特性を定義することによって種のクレードに分類されることがあります:それはラクトースを代謝することができますか?それはクエン酸塩で成長することができますか?それはH₂Sを産生しますか?それは炭水化物を発酵させて酸と気体を作り出しますか?それは嫌気性条件下で成長することができますか?代謝および代謝産物は酵素経路の産物であり、そして酵素は遺伝子にコードされているので、原核生物の代謝能力はそのゲノムを反映しています。地球上には、さまざまなエネルギー源や炭素源、そして原核生物が適応することができるかもしれないさまざまな条件を持つ多くの異なる環境があります。原核生物は、利用可能であるどのようなエネルギー源や炭素源をも利用して、深海の火山孔から南極の氷まで、あらゆる環境で生きることができました。原核生物は、窒素と炭素の循環への関与、酸素の光合成生産、死んだ生物の分解、そして人間を含む多細胞生物内の寄生生物、片利共生生物、または相利共生生物として繁栄することを含む、地球上の多くのニッチを埋めています。原核生物が非常に広範囲の環境を占めることが可能なのは、それらが多様な代謝プロセスを有するからです。
原核生物のニーズ
地球上の多様な環境と生態系は、温度、利用可能な栄養素、酸性度、塩分濃度、酸素の利用可能性、そしてエネルギー源の点で幅広い範囲の条件を持っています。原核生物は、非常に広範な栄養素や環境条件で生活していくためのかなり良い能力を備えています。生きるためには、原核生物はエネルギー源、炭素源、そしていくつかの追加の栄養素を必要とします。
主要栄養素
細胞は、本質的には高分子と水がうまく編成された集合体です。高分子はモノマーと呼ばれる小さな単位の重合によって生成されることを思い出してください。細胞が生命を維持するのに必要なすべての分子を構築するためには、それらは特定の物質を必要とし、それはまとめて栄養素と呼ばれます。原核生物が自然の中で成長するとき、それらは環境から栄養素を得なければなりません。大量に必要とされる栄養素は主要栄養素と呼ばれ、少量または微量が必要とされるものは微量栄養素と呼ばれます。ほんのいくつかの元素が主要栄養素とみなされます — 炭素、水素、酸素、窒素、リン、硫黄です。(これらの元素を覚えるための語呂合わせは頭文字のCHONPSです。)
なぜこれらの主要栄養素は大量に必要なのでしょうか?それらは水を含む細胞中の有機化合物の成分です。炭素はすべての高分子の主要な元素です:炭水化物、タンパク質、核酸、脂質、そして他の多くの化合物。炭素は細胞の組成の約50%を占めています。これとは対照的に、窒素は典型的な細胞の全乾燥重量のわずか12%にすぎません。窒素は、タンパク質、核酸、およびその他の細胞成分の構成要素です。自然で利用可能な窒素の大部分は、大気中の窒素(N₂)または他の無機的な形態のいずれかです。しかしながら、二原子(N₂)窒素は、窒素固定生物と呼ばれる特定の微生物のみによって有機的な形態に変換することができます。水素および酸素も、多くの有機化合物と水の一部です。リンは、ヌクレオチドおよびリン脂質の合成のためにすべての生物によって必要とされます。硫黄は、システインやメチオニンなどのいくつかのアミノ酸の構造の一部であり、いくつかのビタミンや補酵素の中にも存在します。他の重要な主要栄養素はカリウム(K)、マグネシウム(Mg)、カルシウム(Ca)、そしてナトリウム(Na)です。これらの元素は少量が必要とされますが、それらは原核細胞の構造および機能にとって非常に重要です。
微量栄養素
これらの主要栄養素に加えて、原核生物は少量のさまざまな金属元素を必要とします。これらは微量栄養素または微量元素と呼ばれます。たとえば、鉄は電子伝達反応に関与するシトクロムの機能に必要です。原核生物の中には、主に酵素補助因子として、ホウ素(B)、クロム(Cr)、マンガン(Mn)などの他の元素を必要とするものもあります。
原核生物がエネルギーを得る方法
原核生物は、エネルギーを得る方法と有機分子を生産するのに使う炭素源との両方によって分類されます。これらのカテゴリーは表22.3にまとめられています。原核生物は生合成や他の細胞活動に必要なATPを生成するために異なるエネルギー源を使うことができます。光栄養生物(または光栄養性の生物)は、太陽光からエネルギーを得ます。光栄養生物は、クロロフィル、またはいくつかの場合には細菌のロドプシンを使用して光のエネルギーをとらえます。(ロドプシンを使用する光栄養生物は、奇妙なことに、光栄養性ですが、それらは炭素を固定しないので光合成性ではありません。)化学栄養生物(または化学合成生物)は、化学物質からエネルギーを得ます。エネルギー源として有機化合物を使用することができる化学栄養生物は、化学有機栄養生物と呼ばれます。エネルギー源として硫黄または鉄化合物のような無機化合物を使用することができるものは、化学無機栄養生物と呼ばれます。
エネルギー生成経路は、最終電子受容体として酸素を使用する好気性か、または最終電子受容体として単純な無機化合物または有機分子のいずれかを使用する嫌気性のいずれかです。原核生物は、好気呼吸のための大量の酸素が光合成によって生産される以前に、およそ10億年の間地球上に住んでいたために、細菌と古細菌の両方の多くの種は嫌気性であり、それらの代謝活動は後述する炭素と窒素の循環において重要です。
原核生物が炭素を得る方法
原核生物は、さまざまなエネルギー源を使用することができるだけでなく、さまざまな炭素化合物源を使用することもできます。独立栄養原核生物は二酸化炭素から有機分子を合成します。対照的に、従属栄養原核生物は有機化合物から炭素を得ます。原核生物がエネルギーと炭素をどのように取得するかを表す用語を組み合わせると、状況はさらに複雑になります。したがって、光独立栄養生物は太陽光からのエネルギー、および二酸化炭素と水からの炭素を使用するのに対して、化学従属栄養生物は有機化学物質源からエネルギーと炭素の両方を取得します。化学無機独立栄養生物は無機化合物からエネルギーを得て、二酸化炭素から複雑な分子を構築します。最後に、光からエネルギーを得るものの有機化合物から炭素を得る原核生物は、光従属栄養生物です。下の表(表22.3)は原核生物の炭素源とエネルギー源をまとめたものです。
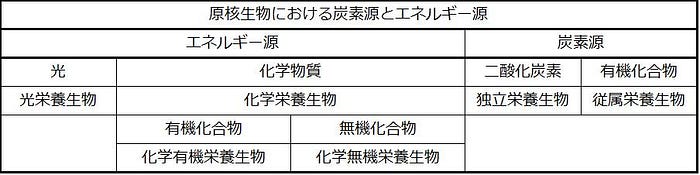
生態系における原核生物の役割
原核生物はいたるところに存在します:それらが存在しないニッチまたは生態系はありません。原核生物はそれらが占める環境において多くの役割を果たします。炭素と窒素の循環でそれらが果たす役割は、地球上の生命にとって不可欠です。さらに、現在の科学的コンセンサスは、代謝的に相互作用する原核生物の生物群集が真核細胞の出現の基盤であったかもしれないことを示唆しています。
原核生物と炭素循環
炭素は最も重要な主要栄養素の1つであり、原核生物は炭素循環において重要な役割を果たしています(図22.18)。炭素循環は、無機化合物から有機化合物へ、そしてまた無機化合物へ戻るという炭素の移動をたどるものです。炭素は地球の主要な蓄積場所、すなわち土地、大気、水中環境、堆積物や岩石、そしてバイオマスを通じて循環します。ある意味では、炭素循環は古代ギリシャの哲学者エンペドクレスによって最初に提案された、火、水、土、そして空気という「四大元素」の役割を反響しています。二酸化炭素は陸上植物および海洋原核生物によって大気から除去され、原核生物、菌類、および動物を含む化学有機栄養生物の呼吸によって大気に戻されます。陸上生態系で最大の炭素の蓄積場所は岩石や堆積物の中ですが、その炭素はすぐには利用できません。
炭素循環の参加者は、有機炭素化合物の生産者、消費者、分解者に大別されます。CO₂から有機炭素化合物を作り出す一次生産者は陸上植物と光合成細菌です。大量の利用可能な炭素は、生きている陸上植物の中に見つけられます。関連する炭素化合物の源は腐植土であり、腐植土は死んだ植物からの有機物質と分解に耐性のある原核生物の混合物です。(ちなみに、「腐植(humus)」という用語は、「人間(human)」という単語の語根です。)動物や他の従属栄養生物などの消費者は、生産者によって生成された有機化合物を使用し、二酸化炭素を大気中に放出します。まとめて分解者と呼ばれる他の細菌および菌類は、動植物およびそれらの有機化合物の分解を行います。大気中の大部分の二酸化炭素は、死んだ動物、植物、腐植土を分解する微生物の呼吸に由来します。
水中環境とその無酸素堆積物では、別の炭素循環が起こります。この場合、循環は一炭素の化合物に基づいています。無酸素堆積物では、原核生物(主に古細菌)がメタン(CH₄)を生成します。このメタンは堆積物の上の区域に移動します。そこは酸素が豊富で、メタンを二酸化炭素に酸化して大気に戻すメタン酸化生物と呼ばれる細菌を支えます。
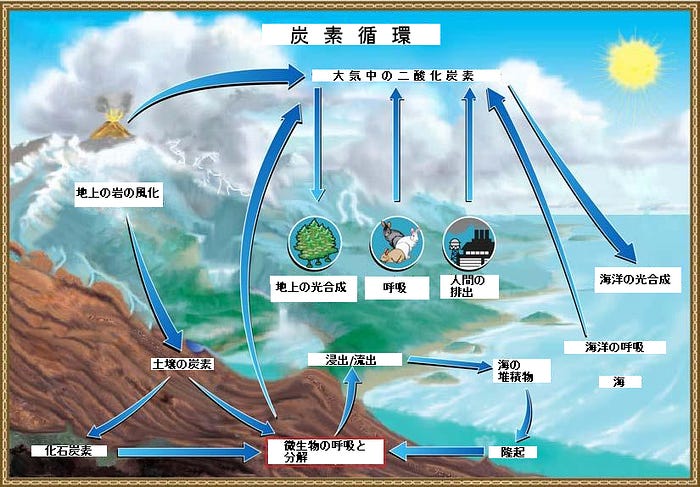
原核生物と窒素循環
窒素は、タンパク質や核酸の主要な成分であるため、生命にとって非常に重要な元素です。それは主要栄養素であり、そして自然の中では、多くのプロセスによって有機化合物からアンモニア、アンモニウムイオン、硝酸塩、亜硝酸塩、および窒素気体にリサイクルされ、そのプロセスの多くは原核生物によってのみ行われます。図22.19に示されるように、原核生物は窒素循環にとっての鍵です。陸上生態系で利用可能な最大の窒素の貯蔵場所は、大気中の気体状窒素(N₂)ですが、この窒素は一次生産者である植物では使用できません。気体状窒素は、窒素固定のプロセスを通じて、アンモニア(NH₃)のようなより容易に利用可能な形態に変換、すなわち「固定」されます。窒素固定細菌には、土壌中のアゾトバクターおよび遍在する光合成シアノバクテリアが含まれます。リゾビウムのようないくつかの窒素固定細菌は、マメ科植物の根の中で共生関係で生きています。アンモニアの別の供給源はアンモニア化であり、これは窒素含有有機化合物の分解中にアンモニアが放出されるプロセスです。アンモニウムイオンは硝化と呼ばれるプロセスでさまざまな種類の細菌によって徐々に酸化されます。硝化プロセスは、アンモニウムから亜硝酸塩(NO₂⁻)への変換で始まり、亜硝酸塩から硝酸塩への変換が続きます。土壌の硝化は、ニトロソモナス属、ニトロバクター属、およびニトロスピラ属に属する細菌によって行われます。土壌中のほとんどの窒素は、アンモニウム(NH₄⁺)または硝酸塩(NO₃⁻)の形をしています。アンモニアと硝酸塩は、植物によって使用されるか、または他の形態に変換されます。
しかしながら、大気中に放出されたアンモニアは、放出された全窒素の15%にすぎません。残りはN₂およびN₂O(亜酸化窒素)として存在します。アンモニアは、いくつかの原核生物によって嫌気的に異化され、最終生成物としてN₂を生じます。脱窒細菌は硝化の過程を逆転させ、土壌の硝酸塩をN₂O、NO、N₂などの気体化合物に還元します。
ビジュアルコネクション

窒素循環についての次の記述のうち、間違っているものはどれですか?
a.窒素固定細菌はマメ科植物の根粒および土壌中に存在する。
b.脱窒細菌は硝酸塩(NO₃⁻)を窒素気体(N₂)に変換する。
c.アンモニア化は、分解する有機化合物からアンモニウムイオン(NH₄⁺)が放出されるプロセスである。
d.硝化は、亜硝酸塩(NO₂⁻)がアンモニウムイオン(NH₄⁺)に変換されるプロセスである。
22.4 | 人間における細菌性疾患
この節が終わるまでに、あなたは次のことができるようになります:
•歴史的に重要な伝染病と流行病を引き起こした細菌性疾患を特定する
•バイオフィルムと食品媒介疾患との関連性を記述する
•抗生物質の過剰使用が、どのようにして「スーパーバグ」を作り出す可能性があるかを説明する
•抗生物質耐性の問題に関してMRSAの重要性を説明する
原核生物にとって、人間は単なる1つの住処の機会かもしれません。残念ながら、いくつかの種が住み着くことは有害な影響を及ぼし、病気を引き起こすことがあります。人間の宿主に害を及ぼす細菌やその他の感染因子は病原体と呼ばれています。壊滅的な病原体媒介性の疾病および疫病は(自然の中ではウイルス性でも細菌性でもあります)、何百万年もの間、人間とその祖先に影響を及ぼしてきました。これらの病気の真の原因は現代の科学的思考が発展するまで理解されておらず、多くの人々はそれらの病気が「超自然的な罰」であると考えていました。苦しんでいる人から離れること、病気の犠牲者の遺体や身に着けていた物を処分すること、そして衛生習慣によって病気になる可能性が減ることを人々が理解したのは、わずかここ数世紀になってからのことです。
疫学者は、病気がどのように伝染し、集団にどのようにして影響を与えるかを研究しています。しばしば彼らは、流行病(ある集団の中で異常に多数の個人が同時に発症する疾患)の過程をたどらなければなりません。これとは対照的に、パンデミックは広い範囲に広がった、通常は世界中で流行しているものです。風土病は、集団の中に常に(通常は低い発生率で)存在する病気です。
細菌性疾患の長い歴史
紀元前3000年にまでさかのぼる感染症についての記録があります。細菌によって引き起こされた多数の重大なパンデミックが数百年にわたって文書化されてきました。最も印象的なパンデミックのいくつかは、都市と国家全体の衰退をもたらしました。
ここ数十年の間に医学研究と治療が進歩したにもかかわらず、21世紀において、感染症は世界の主要な死因の中に残っています。原因となる病原体がある人から別の人に伝染すると、病気は広がります。病原体が病気を引き起こすためには、宿主の体内で繁殖し、何らかの形で宿主に損傷を与えることができなければなりません。
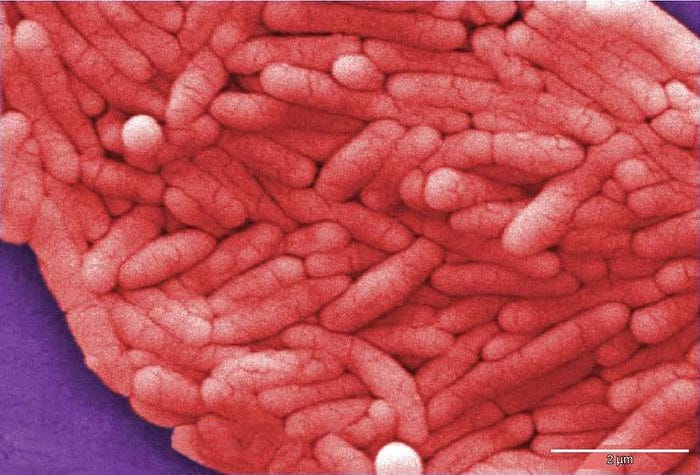
アテナイの疫病
紀元前430年、アテナイの疫病がペロポネソス戦争で戦っていたアテナイ軍の4分の1を殺し、アテナイの支配と権力を弱めました。この疫病は、過密状態のアテナイに住む人々やアテナイに帰るべき船に乗っていた部隊に影響を与えました。近年、アテネ大学の研究者が集団埋葬地から回収した歯からのDNAを使用することができたために、この疫病の原因が特定されたかもしれません。この科学者らは、腸チフスを引き起こす病原性細菌のチフス菌(サルモネラのチフス血清型)(図22.20)からのヌクレオチド配列を同定しました。[3]この疾患は一般に人口過密地域で見られ、記録された歴史を通じて流行を引き起こしてきました。
[3] Papagrigorakis MJ, Synodinos PN, and Yapijakis C. Ancient typhoid epidemic reveals possible ancestral strain of Salmonella enterica serovar Typhi. Infect Genet Evol 7 (2007): 126–7, Epub 2006 Jun.
腺ペスト
541年から750年にかけて、ユスティニアヌスの疫病(恐らくは腺ペストの大流行)は、地中海東部地域の人口の4分の1から2分の1を死亡させました。この流行の間にヨーロッパの人口は50%減少しました。驚くべきことに、腺ペストはヨーロッパを何度も襲うことになります。
腺ペストは細菌エルシニア・ペスティス(Yersinia pestis)によって引き起こされます。腺ペストに起因する最も壊滅的なパンデミックの1つは黒死病(1346年から1361年)でした。それは中国を起源とし、シルクロード(陸と海の貿易ルートのネットワーク)に沿って、常に船の中にいたクマネズミ上に生息するノミによって運ばれて、地中海地域とヨーロッパに広がったと考えられています。黒死病はおそらく、症状の1つである組織の壊死(図22.21c)にちなんで名付けられました。腺ペストの「腺」はリンパ組織における痛みを伴う腫れた領域のことでした。感染した個人の咳やくしゃみによって広がるこの疫病の肺炎型は、人間から人間へと直接広がり、1週間以内に死亡することがあります。この肺炎型はヨーロッパにおける黒死病の急速な拡大の原因でした。黒死病により、世界の人口は推定4億5000万人から約3億5000万~3億7500万人へと減少しました。1600年代半ばには、再び腺ペストがロンドンを襲いました(図22.21)。現代では、毎年約1000~3000件のペストが世界中で発生しています。アメリカ南西部では、プレーリードッグやクロアシイタチなどのげっ歯類に生息するノミが運ぶ、「森林」型のペストに、毎年10~20人が感染しています。抗生物質が登場するより以前に腺ペストに感染することは、ほぼ確実に死亡することを意味していましたが、この細菌は数種類の現代の抗生物質に反応するため、ペストによる死亡率は現在では非常に低くなっています。

学習へのリンク
黒死病(14世紀のヨーロッパにおける腺ペスト)の現代的な理解についてのビデオ(http://openstaxcollege.org/l/black_death)をご覧ください。
新しい集団への病気の移動
人間による探検の悪影響の1つは、病原体をそれ以前にさらされたことがない集団へもたらすことから生じた予想外の「生物学的戦争」でした。何世紀にもわたって、ヨーロッパ人は風土病の感染症に対する遺伝免疫を発展させる傾向がありました。しかし、ヨーロッパの征服者が西半球に達したとき、彼らは病気を引き起こす細菌とウイルスを持ち込み、ネイティブ・アメリカン(ヨーロッパの多くの病気に対して自然な耐性を持っていませんでした)の多くの多様な集団を完全に破壊する流行を引き起こしました。ネイティブ・アメリカンの最大90%がヨーロッパ人の到着後に感染症で死亡したと推定されており、新世界の征服は当然の結論となりました。
新興感染症と再興感染症
特定の病気の分布は動的です。環境、病原体、または宿主集団の変化は、病気の拡大に劇的な影響を及ぼします。世界保健機関(WHO)によると、新興感染症(図22.22)とは、ある集団に初めて出現した、または以前に存在した可能性があるが、発生事例または地理的範囲が急速に増加しているものです。この定義には、以前に制御されていた再興感染症も含まれます。人間に影響を与える最近の新興感染症のおよそ75%は人獣共通感染症です。人獣共通感染症は、主に動物に感染する病気ですが、人間に感染することができるものです。いくつかのものはウイルス起源であり、いくつかのものは細菌起源です。ブルセラ症はいくつかの地域で再興している原核生物による人獣共通感染症の一例であり、そして壊死性筋膜炎(一般に人食いバクテリアとして知られています)は未知の理由からこの80年の間、悪性を増加させ続けています。
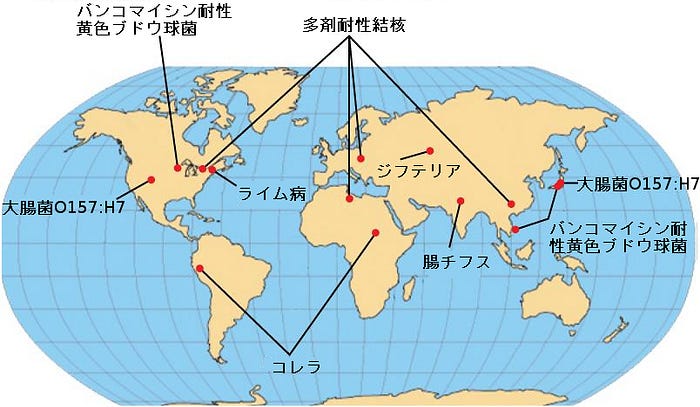
現在の新興感染症の中には、実際には新しいものではなく、過去に壊滅的であった疾患があります(図22.23)。それらは集団を壊滅させ、しばらくの間休眠状態になり、そして時には腺ペストの場合のように以前よりももっと毒性を強めて戻ってきます。結核のような他の病気は根絶されることはありませんでしたが、世界のいくつかの地域では制御下に置かれていました(主に高い割合で免疫的に無防備状態の人々がいる都市部に戻ってくるまでは)。WHOは、世界規模での再興を監視すべき特定の疾患を特定しました。これらの中には、3つのウイルス性疾患(デング熱、黄熱病、およびジカ熱)、および3つの細菌性疾患(ジフテリア、コレラ、および腺ペスト)があります。感染症との闘いにはしばらくの間終わりはありません。
食品媒介疾患
原核生物はいたるところにいます:それらはあらゆる種類の物質の表面に容易にコロニーを形成し、そして食物も例外ではありません。ほとんどの場合、先に私たちが議論したように、原核生物は食品や食品加工機器にバイオフィルムの形でコロニーを形成します。食物摂取に関連した細菌感染の発生は一般的です。食品媒介疾患(一般的に「食中毒」と呼ばれます)は、食物を汚染する病原性の細菌、ウイルス、またはその他の寄生生物の摂取から生じる病気です。米国は食料の供給が世界で最も安全な国の1つですが、米国疾病管理予防センター(CDC)は、「食品媒介疾患によってアメリカ人は毎年7600万人が病気にかかり、30万人以上が入院し、5000人が死亡している」と報告しています。
食品媒介疾患の特徴は時間とともに変化しています。過去において、ボツリヌス中毒症(嫌気性細菌のボツリヌス菌からの毒素によって引き起こされる潜在的に致命的な病気)の散発的な事例について聞くことは比較的一般的でした。この細菌の最も一般的な供給源のいくつかは、非酸性の缶詰食品、自家製のピクルス、および加工肉とソーセージでした。缶、瓶、またはパッケージは、ボツリヌス菌が成長するのに適した嫌気性環境を作り出しました。適切な滅菌と缶詰め手順によって、この病気の発生は減少しました。
人々は食品媒介疾患を動物由来の食品に関連するものとして考える傾向があるかもしれませんが、現在ではほとんどの場合は農産物に関連しています。米国では生のほうれん草、そしてドイツでは芽野菜に関連した深刻な農産物関連の流行があり、これらの種類の流行がより一般的になってきています。2006年の生のホウレンソウでの流行は、細菌である大腸菌の血清型O157:H7によって引き起こされました。ある血清型は、その細胞表面に一連の類似抗原を保有する細菌の1つの株であり、多くの場合、1つの細菌の種にはさまざまな血清型があります。ほとんどの大腸菌は人間にとって特に危険ではありませんが、血清型O157:H7は血性下痢を引き起こすことがあり、潜在的に致命的となることがあります。
あらゆる種類の食品が細菌で汚染される可能性があります。CDCによって報告された最近のサルモネラの発生は、ピーナッツバター、アルファルファの芽、卵などの多様な食品で発生しました。2010年のドイツでの致死的な流行は、芽野菜が大腸菌に汚染されたことによるものです(図22.24)。この流行の原因となった菌株は、以前には他の流行に関与していなかった新しい血清型であることがわかりました。これは、大腸菌が継続的に進化していることを示しています。リステリア・モノサイトゲネス(Listeria monocytogenes)による肉、生のチーズ、冷凍または新鮮な野菜の汚染によるリステリア症の流行は、より頻繁に起こるようになっています。

バイオフィルムと病気
バイオフィルムは破壊するのが非常に難しい微生物群集であることを思い出してください。それらはレジオネラ症、中耳炎(耳の感染症)、そして嚢胞性線維症患者におけるさまざまな感染症のような病気の原因となっています。それらは歯垢を作り出し、カテーテル、人工装具、経皮的および整形外科的機器、コンタクトレンズ、そしてペースメーカーのような内部装置にコロニーを作ります。それらはまた、開放創傷および熱傷組織に形成されます。医療環境では、バイオフィルムは血液透析装置、人工呼吸器、シャント、およびその他の医療器具で成長します。実際のところ、病院で感染したすべての感染症(院内感染症)の65%がバイオフィルムに起因しています。バイオフィルムはまた、食品から感染する病気にも関連しています。なぜならそれらは、野菜の葉や肉の表面、および適切に洗浄されていない食品加工機器の表面にコロニーを形成するためです。
バイオフィルム感染症は徐々に発症し、即時の症状を引き起こさないかもしれません。それらが宿主の防御機構によって解決されることはめったにありません。ひとたびバイオフィルムによる感染症が確立されると、根絶するのは非常に困難です。なぜなら、バイオフィルムは、抗生物質を含む微生物の増殖を制御するために使用されるほとんどの方法に対して耐性がある傾向があるからです。細胞を基質や他のものに付着させるマトリックスは、細胞を抗生物質または薬物から保護します。加えて、バイオフィルムはゆっくり成長するので、それらは細胞成長を妨害する薬剤に対する反応性が低いです。バイオフィルムは、自由生活性またはプランクトン性の同じ細菌を殺すのに使用される抗生物質濃度の最大1000倍の濃度にまで耐性があることが報告されています。そこまで大きな抗生物質の量だと、患者に害を及ぼすことになります。したがって、科学者たちはバイオフィルムを取り除くための新しい方法に取り組んでいます。
抗生物質:私たちは危機に直面しているのでしょうか?
抗生物質(アンチバイオティック)という言葉は、「反対」を意味するアンチと、「生命」を意味するバイオというギリシャ語に由来します。抗生物質は、微生物によって、または合成によって生産される化学物質であって、他の生物に敵対的であったりその増殖を防いだりするものです。今日のメディアはしばしば抗生物質の危機についての懸念を取り上げています。過去に細菌感染を容易に治療した抗生物質は時代遅れになりつつあるのでしょうか?新しい「スーパーバグ」 — 私たちの抗生物質の蓄積に対してより耐性を持つように進化した細菌 — がいるのでしょうか?これは抗生物質の終わりの始まりですか?これらすべての質問は、医療界に挑戦を突き付けています。
細菌の抗生物質への耐性の主な原因の1つは、抗生物質への過剰な暴露です。抗生物質の無思慮かつ過度の使用は、耐性型の細菌の自然選択をもたらしました。抗生物質は感染する細菌の大部分を殺しますが、それゆえに耐性型だけが残ります。これらの耐性型は増殖し、その結果、非耐性型よりも耐性型の割合が増加します。耐性遺伝子の子孫への伝達に加えて、プラスミド上での耐性遺伝子の横方向伝播は細菌集団にわたってこれらの遺伝子を急速に広げることができます。抗生物質の主な誤用は、風邪やインフルエンザなどのウイルス感染症の患者に行うものであり、それらに対しては抗生物質は役に立ちません。もう1つの問題は家畜における抗生物質の過剰使用です。動物飼料中に抗生物質を日常的に使用することも、細菌の耐性を促進します。米国では、生産された抗生物質の70%が動物に与えられています。これらの抗生物質は家畜に対して低用量で投与され、それは耐性発生の可能性を最大にし、そしてこれらの耐性細菌は容易に人間へと移されます。
学習へのリンク
家畜への日常的な抗生物質投与と抗生物質耐性細菌の問題に関する最近のニュースレポート(http://openstaxcollege.org/l/antibiotics)をご覧ください。
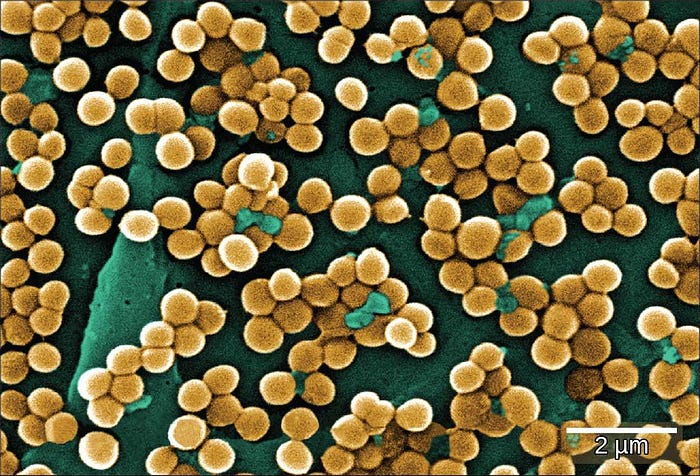
スーパーバグの1つ:MRSA
抗生物質の不用意な使用は、耐性細菌の個体群の拡大への道を開きました。たとえば、黄色ブドウ球菌(Staphylococcus aureus)は、しばしば「スタフ(staph)」と呼ばれる人体に住むことができる一般的な細菌であり、通常は抗生物質で簡単に治療できます。しかしながら、非常に危険な菌株であるメチシリン耐性黄色ブドウ球菌(MRSA)が、過去数年にわたってニュースになっています(図22.25)。この株は、メチシリン、アモキシシリン、ペニシリン、およびオキサシリンを含む多くの一般的に使用されている抗生物質に対して耐性があります。MRSAは皮膚の感染症を引き起こすことがありますが、それはまた血流、肺、尿路、または損傷部位にも感染することがあります。MRSA感染症は医療施設の人々の間で一般的ですが、それは入院していないものの人口が密集している場所に住んだり働いたりしている健康な人々(軍人や囚人のような)にも発生しています。研究者らは、この後者のMRSAの発生源が、医療施設に滞在する人々よりもはるかに若い人々をターゲットにしていることについて懸念を表明しています。ジャーナル・オブ・ジ・アメリカン・メディカル・アソシエーション(米国医師会ジャーナル)では、医療施設でMRSAに罹患している人の平均年齢は68歳であるのに対し、「コミュニティー関連MRSA」(CA-MRSA)の人の平均年齢は23歳であることが報告されています。[4]
[4] Naimi, TS, LeDell, KH, Como-Sabetti, K, et al. Comparison of community- and health care-associated methicillin-resistant Staphylococcus aureus infection. JAMA 290 (2003): 2976–84, doi: 10.1001/jama.290.22.2976.
要約すると、医学界は抗生物質の危機に直面しています。一部の科学者たちは、私たちが何年にもわたって抗生物質によって細菌感染から保護されてきた後、私たちは再び単純な細菌感染が人類集団を荒廃させるおそれのある時代に戻りつつある、と考えています。研究者たちは新しい抗生物質を開発していますが、効果的で承認された薬を生み出すには長年の研究と臨床試験に加えて数百万ドルの財政的投資が必要とされます。
キャリアへのつながり
疫学者
疫学とは、ある集団における健康と病気の発生、分布、および決定要因の研究のことです。したがって、それは公衆衛生の一部です。疫学者は、人間の集団や環境内での病気の頻度と分布を研究しています。
疫学者は、特定の疾患についてのデータを収集し、その広がりを追跡して、最初の感染様式を特定します。彼らは時には歴史家と密接に協力して、ある病気が地理的にそして時間の経過とともにどのように進行したかを理解しようと試み、病原体の自然史を調査します。彼らは、臨床記録、患者へのインタビュー、観察、およびその他の利用可能なすべての手段からの情報を収集します。その情報は、ワクチン接種などの戦略を立案し(図22.26)、病気の発生を減らしたり流行を防いだりするための公衆衛生政策を設計することに使用されます。疫学者はまた、大流行が発生した場合には、それを管理するための早急な対策を推奨するために、迅速な調査を行います。
疫学者は学士号に加えて公衆衛生学の修士号(MPH)を取得しています。多くの疫学者は医師でもあり(そして医学博士号(M.D.)または外科医学博士号(D.O.)を取得しています)、あるいは彼らは生物学や微生物学などの関連分野で博士号(Ph.D.)を取得しています。

22.5 | 有益な原核生物
この節が終わるまでに、あなたは次のことができるようになります:
•窒素固定の必要性とその達成方法を説明する
•私たちの皮膚や消化管にコロニーを形成する細菌の有益な効果を記述する
•食品加工中に使用される原核生物を特定する
•バイオレメディエーションにおける原核生物の使用について記述する
幸いなことに、病原性の原核生物はごくわずかです!原核生物はまた、有益であるような多くの方法で人間や他の生物と相互作用します。たとえば、原核生物は炭素と窒素の循環の主要な参加者です。それらは人間や他の動物の消化管で栄養素を作り出すか、または処理を行います。原核生物は、いくつかの人間の食品の生産に使用されており、また有害物質の分解のために採用されています。実際のところ、私たちの生活は原核生物なしには不可能です。
細菌と真核生物の協力:窒素固定
窒素は、それぞれ核酸とタンパク質の基礎的要素であるヌクレオチドとアミノ酸の一部であるため、生物にとって非常に重要な元素です。窒素は通常、陸上生態系で最も限られた元素であり、大気中の窒素N₂が利用可能な窒素の最大の蓄積場所を提供しています。しかしながら、真核生物は、高分子を合成するために大気中の気体の窒素を使用することはできません。幸いなことに、窒素は「固定」することができます。つまりそれは、生物的にも非生物的にも、より利用しやすい形のアンモニア(NH₃)に変換することができます。
非生物的な窒素固定は、落雷などの物理的過程、または工業的過程の結果として起こります。生物的な窒素固定(BNF)は、原核生物:土壌細菌、シアノバクテリア、およびフランキア属(ハンノキ、ヤマモモ、スウィートファーンといった、放線菌と共生する植物と相互作用する糸状菌)によって独占的に行われます。BNFは光合成に次いで地球上で最も重要な生物学的プロセスです。以下の窒素固定の方程式の全体は、一連の酸化還元反応を表しています(Piは無機リン酸塩を表します)。
N₂ + 16ATP + 8e⁻ + 8H⁺ → 2NH₃ + 16ADP + 16Pi + H₂
BNFによる総固定窒素量は年間約1億~1億8000万トンで、農業で使用される窒素の約65%を占めています。
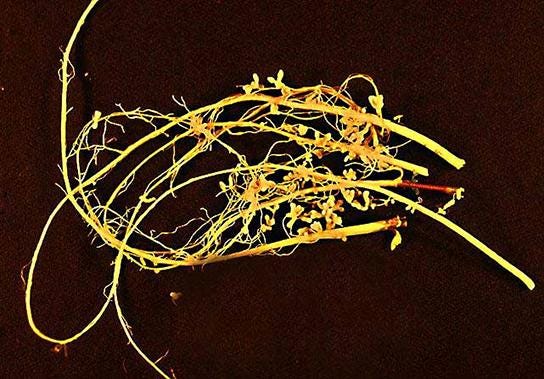
シアノバクテリアは水中環境で最も重要な窒素固定生物です。土壌中では、クロストリジウム属およびアゾトバクター属のメンバーが、自由生活性の窒素固定細菌の例です。他の細菌はマメ科植物と共生し、最も重要な固定窒素源を提供します。共生生物は、土壌中で自由生活性の生物よりも10倍も多く窒素を固定することができます。まとめて根粒菌と呼ばれる土壌細菌は、マメ科植物と共生的に相互作用して、窒素固定が起こる特殊な構造である根粒を形成することができます(図22.27)。窒素を固定する酵素であるニトロゲナーゼは酸素によって不活性化されるので、根粒は窒素固定を行うための無酸素領域を提供します。酸素はレグヘモグロビンと呼ばれる植物ヘモグロビンの一種によって隔離されます。レグヘモグロビンはニトロゲナーゼを保護するものの、呼吸活動を支えるのに十分な酸素を放出します。
共生窒素固定は天然で安価な植物肥料を提供します。それは大気中の窒素を、植物によって容易に使用可能であるアンモニアに還元します。マメ科植物の使用は化学肥料に代わる優れた方法であり、化学物質の使用を最小限に抑え、天然資源を節約しようとする持続可能な農業にとって特に興味深いものです。共生窒素固定を通じて、植物は無限の窒素源、すなわち大気を使うことからの利益を得ます。細菌は植物からの光合成産物(光合成の間に生産される炭水化物)を使用すること、そして保護されたニッチを持つことから利益を得ます。さらに、土壌は自然に施肥されることから利益を得ます。したがって、生物肥料としての根粒菌の使用は持続可能な実践となります。
なぜマメ類はそんなにも重要なのでしょうか?大豆のように、いくつかのものは農業用タンパク質の主要な供給源です。人間によって消費される最も重要なマメ類のいくつかは、大豆、ピーナッツ、エンドウマメ、ヒヨコマメ、および小豆です。アルファルファなどの他のマメ科植物は、牛の餌に使われています。
日常へのつながり
人体の微生物
私たちの皮膚や胃腸管に生息する片利共生細菌は、私たちにとってたくさんの良いことをしています。それらは私たちを病原体から守り、私たちが食物を消化するのを助け、そしてビタミンや他の栄養素の一部を生産します。これらの活動は長い間知られています。より最近では、科学者たちはこれらの細菌が私たちの気分を調節し、私たちの活動レベルに影響を与え、さらには私たちの食べ物の選択と吸収パターンに影響を与えることによって体重を制御するのを助けることまでしているのかもしれないという証拠を集めています。ヒト・微生物叢プロジェクト(Human Microbiome Project)は、私たちがこれらの機能をよりよく理解できるように、正常な細菌(および古細菌)の目録を作成するプロセスを開始しました。
私たちの正常細菌叢の特に魅力的な例は、私たちの消化器系に関連しています。高用量の抗生物質を服用する人は、正常な腸内細菌の多くを失う傾向があり、クロストリジウム・ディフィシル(Clostridium difficile)と呼ばれる天然の抗生物質耐性種が過剰に増殖し、重度の胃腸の問題(特に慢性下痢)を引き起こすことになります(図22.28)。明らかに、この問題を抗生物質で治療しようとすることは、それを悪化させるだけです。しかしながらそれは、正常な腸内微生物群集を再確立するために患者に対して健康なドナーからの便移植を与えることによってうまく治療することができます。この技術の安全性と有効性を確保するために臨床試験が進行中です。

科学者たちはまた、私たちの腸管から特定の重要な微生物が欠如していると、さまざまな問題を引き起こすことがあることを発見しつつあります。これは、免疫システムの適切な機能に関して特に当てはまるようです。これらの微生物の不在がアレルギーといくつかの自己免疫疾患の発症に重要な貢献をしていることを示唆する興味深い発見があります。ある種の微生物を私たちの内部の生態系に加えることがこれらの問題の治療やある種の自閉症の治療に役立つかどうかをテストする研究が現在進行中です。
初期のバイオテクノロジー:チーズ、パン、ワイン、ビール、そしてヨーグルト
国際連合の生物の多様性に関する条約によれば、バイオテクノロジーとは「物又は方法を特定の用途のために作り出し又は改変するため、生物システム、生物又はその派生物を利用する応用技術」です。[5]「特定の用途」の概念は、ある種の商業的な応用も含みます。遺伝子工学、人為選択、抗生物質の生産、細胞培養はバイオテクノロジーにおける現在の研究テーマであり、後の章で説明されるでしょう。しかしながら、バイオテクノロジーという用語が造られる前でも、人間は原核生物を使っていました。この初期のバイオテクノロジーの産物は、チーズ、パン、ワイン、ビール、ヨーグルトといったよく知られたものであり、これらは細菌と、酵母や菌類などの他の微生物との両方を使用しています(図22.29)。
[5] http://www.cbd.int/convention/articles/?a=cbd-02 (http://openstax.org/l/UN_convention) , United Nations Convention on Biological Diversity: Article 2: Use of Terms.

チーズの生産は、人間が動物を飼育し、その乳を加工するようになった4000年から7000年前に始まりました。この場合の発酵は栄養素を保存します。乳は比較的早く腐敗しますが、チーズとして加工するとより安定します。ビールに関しては、醸造の最も古い記録は約6000年前であり、シュメール文化の不可欠な部分でした。証拠は、シュメール人が偶然に発酵を発見したことを示しています。ワインは約4500年の間生産されており、そして、ヨーグルトのような発酵乳製品は少なくとも4000年の間存在しているということが証拠から示唆されています。
私たちの惑星を浄化するために原核生物を用いる:バイオレメディエーション
微生物バイオレメディエーションは、汚染物質を除去するために原核生物(または微生物代謝)を使用することです。バイオレメディエーションは、土壌から地下水および地下へと浸出する農業用化学物質(たとえば、殺虫剤、肥料)を除去するために使用されてきました。セレンやヒ素化合物などの特定の有毒な金属や酸化物もバイオレメディエーションによって水から除去することができます。SeO₄⁻²からSeO₃⁻²へ、そしてSe⁰(金属セレン)への還元は、水からセレンイオンを除去するために使われる方法です。水銀(Hg)はバイオレメディエーションによって環境から除去することができる有毒な金属の一例です。水銀はいくつかの農薬の有効成分として産業界で使用されており、また、電池製造などの特定の行程の副生成物でもあります。メチル水銀は通常、自然環境では非常に低濃度で存在していますが、生体組織に蓄積するため、非常に有毒です。いくつかの種の細菌は、有毒な水銀を無毒な形態に生体内変換することができます。緑膿菌などのこれらの細菌は、Hg⁺²を人間に対して無毒なHg⁰に変換することができます。
バイオレメディエーション目的のための原核生物の使用において最も有用で興味深い例の1つは、流出油の浄化です。石油バイオレメディエーションに対する原核生物の意義は、アラスカのエクソン・ヴァルディーズ号原油流出事故(1989年)(図22.30)、スペインのプレスティージ号重油流出事故(2002年)、レバノンの発電所から地中海への流出(2006年)、そしてより最近ではメキシコ湾でのBPの原油流出事故(2010年)など、近年のいくつかの油流出事故で証明されています。海上での油流出の場合、流出前の海には油を消費する細菌がいるため、持続的な自然のバイオレメディエーションが起こる傾向があります。これらの天然に存在する油分解細菌に加えて、人間は、同じ能力を持つものの、処理可能な炭化水素化合物の種類と効率が向上した細菌を選択して操作します。バイオレメディエーションは細菌の増殖を助ける無機栄養素の添加によって強化されます。
いくつかの炭化水素分解細菌は油滴中の炭化水素を食べ、その炭化水素をより小さなサブユニットに分解します。アルカニボラックス・ボルクメンシス(Alcanivorax borkumensis)などの一部の種は、油を可溶化する(水溶性にする)界面活性剤を生成する一方、他の細菌は油を二酸化炭素に分解します。理想的な条件下では、油中の不揮発性成分の最大80%が流出から1年以内に分解されることがあると報告されています。芳香族鎖および高度に分岐した炭化水素鎖を含む油の他の部分は除去することがより困難であり、そして長期間環境中に留まります。

重要用語
好酸性生物:最適な増殖pHが3以下の生物
好アルカリ性生物:最適な増殖pHが9以上の生物
アンモニア化:窒素含有有機化合物の分解中にアンモニアが放出されるプロセス
嫌気性:酸素なしで成長する生物を指す
無酸素:酸素なしで
抗生物質:低濃度で原核生物の増殖に拮抗する生物学的物質
バイオフィルム:ゴム状の質感のある基質によってまとめられた微生物群集
生物的な窒素固定:原核生物によって独占的に行われる大気中の窒素のアンモニアへの変換
バイオレメディエーション:汚染物質を除去するための微生物代謝の使用
バイオテクノロジー:生物、生物システム、またはそれらの派生物を使用して他の生産物を生産または改変する、あらゆる技術的応用
黒死病:細菌エルシニア・ペスティスによって引き起こされる腺ペストの流行であると考えられている壊滅的なパンデミック
ボツリヌス中毒症:嫌気性細菌のボツリヌス菌の毒素によって引き起こされる疾患
CA-MRSA:病院ではなくコミュニティーの中で罹患したMRSA
莢膜:原核生物が表面に付着すること、そしてそれを脱水から保護することを可能にする外部構造
化学栄養生物:化学物質からエネルギーを得る生物
接合:原核生物が性繊毛を使用してDNAをある個体から別の個体に移動させるプロセス
シアノバクテリア:初期の光栄養生物から進化して大気を酸素化した細菌。藍藻としても知られる
分解者:死んだ生物を分解する生物
脱窒:土壌からの硝酸塩をN₂O、NO、N₂などの気体状窒素化合物へ変換すること
新興感染症:ある集団で初めて出現する疾患、または発生事例または地理的範囲が増加している疾患
風土病:ある集団の中に、通常は低い発生率で、絶えず存在する疾患
伝染病:ある集団内の異常に多数の個人に同時に発生する疾患
好極限性生物:極端な、または過酷な条件下で成長する生物
食品媒介疾患:汚染された食物、または食物を汚染する病原細菌、ウイルス、その他の寄生生物の消費から生じるあらゆる病気
グラム陰性:細胞壁がペプチドグリカンをほとんど含まないが、外膜を持つ細菌
グラム陽性:細胞壁に主にペプチドグリカンを含む細菌
好塩性生物:塩濃度0.2M以上を必要とする生物
熱水噴出孔:地熱によって加熱された水を放出する地球の表面の裂け目
超好熱性生物:80~122°Cの温度で成長する生物
微生物マット:細菌や古細菌を含むことのある原核生物の多層シート
MRSA(メチシリン耐性黄色ブドウ球菌):複数の抗生物質に耐性のある非常に危険な黄色ブドウ球菌株
硝化:アンモニウムを亜硝酸塩と土壌中の硝酸塩へ変換すること
窒素固定:気体状窒素をアンモニアのようなより容易に利用可能な形態に変換、または「固定」するプロセス
根粒:植物と土壌細菌との間の共生的相互作用から生じ、窒素固定の部位であるような特定の植物(マメ科植物)の根の新奇な構造
栄養素:炭素や窒素などの成長に不可欠な物質
高濃性生物:高糖濃度で成長する生物
パンデミック:広範囲に及ぶ、通常は世界中での流行病
ペプチドグリカン:非天然ペプチドによって架橋されている多糖鎖からなる材料
光栄養生物:太陽エネルギーを化学エネルギーに変換することによって自身の食物を作ることができる生物
性繊毛:他の原核生物を含む表面への付着に使用される、いくつかの原核生物の表面の付属器官
疑似ペプチドグリカン:形態はペプチドグリカンに似ているが異なる糖を含む古細菌の細胞壁の成分
好冷性生物:-15°C以下の温度で成長する生物
放射線耐性:高レベルの放射線の中で成長する生物
蘇生:VBNC状態にある原核生物が生存可能な状態へ戻るプロセス
S層:古細菌や細菌の細胞壁の外側に存在する表層タンパク質
血清型:その細胞表面に一連の類似した抗原を保有する細菌の株で、多くの場合1つの細菌の種に含まれるもの
ストロマトライト:微生物マット中の原核生物による鉱物の析出によって形成された層状の堆積構造
テイコ酸:グラム陽性菌の細胞壁と関連するポリマー
好熱性生物:60~80°Cの温度で生活する生物
形質導入:バクテリオファージがある原核生物から別の原核生物へとDNAを移動させるプロセス
形質転換:原核生物が他の原核生物によって環境中にこぼされたDNAを取り込むプロセス
生存可能だが培養不可能(VBNC)状態:環境ストレス条件に直面している細菌の生存機構
人獣共通感染症:主に動物に感染する疾患で、人間に感染するもの
この章のまとめ
22.1 | 原核生物の多様性
原核生物は植物や動物が出現する以前に何十億年もの間存在していました。温泉と熱水噴出孔は生命が始まった環境であったかもしれません。微生物マットは地球上で最も初期の生命の形態を表すと考えられています。微生物マットは、主として湿った表面の上にある、異なる種類の材料の間の界面で成長する原核生物の多層シートです。化石化した微生物マットはストロマトライトと呼ばれ、原核生物による鉱物の析出によって形成された積層の有機堆積構造からなります。それらは地球上での最も初期の生命の化石記録を表しています。
最初の20億年の間、大気は無酸素で、嫌気性生物だけが生きることができました。シアノバクテリアは初期の光栄養生物から進化し、大気の酸素化を始めました。酸素濃度の増加は他の形態の生命の進化を可能にしました。
細菌と古細菌は、事実上あらゆる環境で増殖します。極端な条件下で生き残るものは、好極限性生物(極限環境を愛するもの)と呼ばれます。いくつかの原核生物は実験室の環境で増殖することはできないものの、死んでもいません。それらは生存可能だが培養不可能(VBNC)状態にあります。原核生物が環境ストレス要因に反応して休眠状態に入ると、VBNC状態が発生します。ほとんどの原核生物はコロニーを作るものであり、相互作用が行われる生物群集に住むことを好みます。バイオフィルムとは、ゴム状の質感のある基質にまとめられた微生物群集のことです。
22.2 | 原核生物の構造:細菌と古細菌
原核生物(古細菌と細菌のドメイン)は、核を持たない単細胞生物です。それらは、細胞の核様体領域の中に単一の環状DNAを有します。ほとんどの原核生物は、原形質膜の境界の外側にある細胞壁を持っています。原核生物の中には、莢膜、鞭毛、および性繊毛などの追加の構造を持つものがあります。細菌と古細菌は、それらの細胞膜の脂質組成と細胞壁の特徴において異なります。古細菌の膜では、脂肪酸ではなくフィタニル単位がグリセロールに結合しています。いくつかの古細菌の膜は、二重層ではなく脂質単層です。
細胞壁は細胞膜の外側に位置し、浸透圧溶解を防ぎます。細胞壁の化学組成は種によって異なります。細菌の細胞壁はペプチドグリカンを含みます。古細菌の細胞壁はペプチドグリカンを有しませんが、それらは疑似ペプチドグリカン、多糖類、糖タンパク質、またはタンパク質をベースとした細胞壁を持つことがあります。細菌は、グラム染色反応に基づいて、グラム陽性とグラム陰性の2つの主要なグループに分けることができます。グラム陽性生物は、テイコ酸で強化された厚いペプチドグリカン層を有します。グラム陰性生物は、薄い細胞壁と、リポ多糖類およびリポタンパク質を含む外側エンベロープとを有します。
原核生物は、3つのメカニズムによって、ある細胞から別の細胞へとDNAを移転させることができます:すなわち、形質転換(環境DNAの取り込み)、形質導入(ウイルスを介したゲノムDNAの移転)、および接合(直接の細胞接触によるDNAの移転)です。
22.3 | 原核生物の代謝
原核生物は地球の最も古い生きた住民であり、また、最も代謝的に多様です。それらはさまざまなエネルギー源と炭素源、さまざまな温度、pH、圧力、酸素と水の利用可能性のある多くの異なる環境で繁栄します。大量に必要とされる栄養素は主要栄養素と呼ばれる一方で、微量が必要とされる栄養素は微量栄養素または微量元素と呼ばれます。主要栄養素には、C、H、O、N、P、S、K、Mg、Ca、およびNaが含まれます。これらの主要栄養素に加えて、原核生物は成長と酵素機能のためにさまざまな金属元素を必要とします。原核生物は、より小さな分子から高分子を組み立てるために異なるエネルギー源を使用します。光栄養生物は太陽光からエネルギーを得ますが、化学栄養生物は化学物質からエネルギーを得ます。エネルギー産生経路は、好気的または嫌気的のいずれかであり得ます。
原核生物は炭素と窒素の循環で役割を果たしています。生産者は大気から二酸化炭素をつかまえ、それを有機化合物に変換します。消費者(動物および他の化学有機栄養生物)は、生産者によって生成された有機化合物を使用し、呼吸によって二酸化炭素を大気中に放出します。二酸化炭素は死んだ生物の微生物分解者によっても大気に戻されます。窒素はまた、有機化合物からアンモニア、アンモニウムイオン、亜硝酸塩、硝酸塩、および窒素気体へと、生物の内外を循環します。原核生物は、これらの変換のほとんどにおいて不可欠なものです。気体状窒素は窒素固定によりアンモニアに変換されます。アンモニアは、いくつかの原核生物によって嫌気的に異化され、最終生成物としてN₂を生じます。硝化はアンモニウムの亜硝酸塩への変換です。土壌中の硝化は細菌によって行われます。脱窒も細菌によって行われ、土壌からの硝酸塩をN₂O、NO、N₂などの気体状の窒素化合物に変換します。
22.4 | 人間における細菌性疾患
いくつかの原核生物は人間にとっての病原体です。壊滅的な病気や疫病は、古くから私たちの間で存在し続けており、世界中で主要な死因のひとつとして残っています。新興感染症は、発生事例または地理的範囲が急速に増加しているものです。それらは、新たな疾患または再興感染症(以前は抑制されていた)であり得ます。ブルセラ症など、人間に影響を与える多くの新たな病気は動物に由来します(人獣共通感染症)。WHOによって監視のために最近指定された細菌性の再興感染症のグループには、腺ペスト、ジフテリア、およびコレラが含まれます。食品媒介疾患は、食品、病原菌、ウイルス、または寄生生物で汚染された食品の消費に起因します。
レジオネラ症、中耳炎、および嚢胞性線維症患者の感染症など、いくつかの細菌性感染症はバイオフィルムに関連しています。バイオフィルムは、歯垢のように人間の組織の上で成長したり、医療機器にコロニーを作ったり、食品や食品加工機器の表面で成長することによって感染や食品媒介疾患を引き起こしたりすることがあります。バイオフィルムは、微生物の増殖を制御するために使用されるほとんどの方法に対して耐性があります。抗生物質の過剰使用は、時間がたつにつれて耐性型の細菌が選択されてきたため、世界的に大きな問題となっています。非常に危険な菌株であるメチシリン耐性黄色ブドウ球菌(MRSA)は、最近世界中で大混乱を引き起こしています。
22.5 | 有益な原核生物
病原体はすべての原核生物のうちのごく小さな割合にすぎません。実際のところ、原核生物は人間や他の生物にとって不可欠なサービスを提供しています。窒素(真核生物は豊富な大気形態では使用できません)は、生物的または非生物的のいずれかで「固定」、すなわちアンモニア(NH₃)に変換することができます。生物的窒素固定(BNF)は、原核生物によって独占的に行われており、地球上で2番目に重要な生物学的プロセスを構成しています。一部の陸上の窒素は自由生活性の細菌によって固定されていますが、ほとんどのBNFは土壌根粒菌とマメ科植物の根との間の共生的相互作用に由来します。
人間の生活は、環境中のものと私たちを住処とする種との両方の微生物の働きによってのみ可能です。内部的には、それらは私たちが食物を消化するのを助け、私たちにとって不可欠な栄養素を生産し、私たちを病原性微生物から守り、そして私たちの免疫系が正しく機能するよう訓練するのを助けます。
微生物バイオレメディエーションとは、汚染物質を除去するために微生物の代謝を使用することです。バイオレメディエーションは、土壌から地下水や地下に浸出する農業用化学物質を除去するために使用されてきました。セレンやヒ素化合物などの有毒な金属や酸化物もバイオレメディエーションによって除去することができます。バイオレメディエーション目的のための原核生物の使用においておそらく最も有用で興味深い例の1つは流出油の清掃です。
ビジュアルコネクション問題
1.図22.8 | 自由浮遊性の細菌と比較して、バイオフィルム内の細菌はしばしば抗生物質および洗剤に対する耐性の増加を示します。あなたはなぜそうなのだと思いますか?
2.図22.16 | 次の記述のうち、正しいものはどれですか?
a.グラム陽性菌は、リポテイコ酸によって細胞膜に固定された単一の細胞壁を有する。
b.ポリンはグラム陽性菌とグラム陰性菌の両方への物質の侵入を可能にする。
c.グラム陰性菌の細胞壁は厚く、グラム陽性菌の細胞壁は薄い。
d.グラム陰性菌はペプチドグリカンからなる細胞壁を有するのに対し、グラム陽性菌はリポテイコ酸からなる細胞壁を有する。
3.図22.19 | 窒素循環についての次の記述のうち、間違っているものはどれですか?
a.窒素固定細菌はマメ科植物の根粒および土壌中に存在する。
b.脱窒細菌は硝酸塩(NO₃⁻)を窒素気体(N₂)に変換する。
c.アンモニア化は、分解する有機化合物からアンモニウムイオン(NH₄⁺)が放出されるプロセスである。
d.硝化は、亜硝酸塩(NO₂⁻)がアンモニウムイオン(NH₄⁺)に変換されるプロセスである。
レビュー問題
4.地球上での最初の生命の形態は_________と考えられていました。
a.単細胞の植物
b.原核生物
c.昆虫
d.恐竜などの大型動物
5.微生物マットは、__________。
a.地球上で最も初期の生命の形態である
b.熱水噴出孔からエネルギーと食物を得ていた
c.ほとんどが細菌であるが古細菌も含んでいた原核生物の多層シートである
d.上記のすべて
6.大気を酸素化した最初の生物は、__________です。
a.シアノバクテリア
b.光栄養性生物
c.嫌気性生物
d.上記のすべて
7.好塩性生物は、________を必要とする生物です。
a.塩分濃度0.2M以上
b.高い糖濃度
c.ハロゲンの添加
d.上記のすべて
8.科学研究室で培養される最初の原核生物の多くは、人間または動物の病原体でした。なぜこれらの種は非病原性の原核生物よりも容易に培養されるのでしょうか?
a.病原性の原核生物は、非病原性の原核生物よりも強固だから。
b.非病原性の原核生物は、それらの増殖培地においてより多くの栄養を必要とするから。
c.病原性の原核生物のために必要な培養条件のほとんどは推測できたから。
d.病原性の細菌は遊離細菌として増殖することができるが、非病原性の細菌は大きなコロニーの一部として増殖するだけだから。
9.膜で包まれた核の存在は、________にとっての特徴です。
a.原核細胞
b.真核細胞
c.全ての細胞
d.ウイルス
10.次のうち、原核細胞で構成されているものはどれですか?
a.細菌と菌類
b.古細菌と菌類
c.原生生物と動物
d.細菌と古細菌
11.細胞壁は、________。
a.細胞膜の内部にある
b.細胞膜の外部にある
c.細胞膜の一部である
d.特定の細胞に応じて、内部または外部にある
12.極端な環境で最も発見される可能性が高い生物は________です。
a.菌類
b.細菌
c.ウイルス
d.古細菌
13.原核生物は細胞の_______の違いによって、グラム陽性またはグラム陰性として染色されます。
a.壁
b.細胞質
c.核
d.染色体
14.疑似ペプチドグリカンは、________の壁の特徴です。
a.真核細胞
b.細菌の原核細胞
c.古細菌の原核細胞
d.細菌および古細菌の原核細胞
15.リポ多糖層(LPS)は、________の壁の特徴です。
a.古細菌の細胞
b.グラム陰性菌
c.細菌の原核細胞
d.真核細胞
16.次の元素のうち微量栄養素でないものはどれですか?
a.ホウ素
b.カルシウム
c.クロム
d.マンガン
17.化学物質からエネルギーを得る原核生物は、_____と呼ばれています。
a.光栄養生物
b.独立栄養生物
c.化学栄養生物
d.無機栄養生物
18.アンモニア化は、_____プロセスです。
a.窒素含有有機化合物の分解中にアンモニアが放出される
b.アンモニウムが土壌中で亜硝酸塩と硝酸塩に変換される
c.土壌からの硝酸塩が、NO、N₂O、N₂などの気体状の窒素化合物に変換される
d.気体状の窒素が固定されてアンモニアを生成する
19.植物は大気中の二酸化炭素を使用しているため、_____と呼ばれています。
a.消費者
b.生産者
c.分解者
d.炭素固定者
20.シアノバクテリアは、光合成を通じて太陽からのエネルギーを利用し、水を酸化してエネルギー生成のための電子を供給します。したがって、私たちはシアノバクテリアを_________として分類します。
a.光無機栄養生物
b.光独立栄養生物
c.化学無機独立栄養生物
d.化学有機栄養生物
21.集団の中に絶えず存在している病気は、_____と呼ばれています。
a.パンデミック
b.流行病
c.風土病
d.再興感染症
22.バイオフィルムについての記述のうち、不正確なものはどれですか?
a.バイオフィルムは、嚢胞性線維症などの疾患の原因と考えられている。
b.バイオフィルムは歯垢を形成し、そしてカテーテルや人工装具にコロニーを作る。
c.バイオフィルムは、開放創傷および熱傷組織にコロニーを作る。
d.すべての記述が不正確である。
23.これらの記述のうち、正しいものはどれですか?
a.抗生物質は、原核生物の増殖に拮抗する、生物によって産生されるあらゆる物質である。
b.抗生物質は、他のウイルスの増殖に拮抗する、原核生物によって産生されるあらゆる物質である。
c.抗生物質は、真核細胞の増殖に拮抗する、原核生物によって産生されるあらゆる物質である。
d.抗生物質は、同じ原核生物の増殖を妨げる、原核生物によって産生されるあらゆる物質である。
24.イングランドに住むある人が、ニューメキシコ州への訪問から戻った直後に、発熱とリンパ節の腫れを伴って診療所を訪れました。医師はどの細菌についてこの患者を検査するべきですか?
a.サルモネラ菌
b.ボレリア・ブルグドルフェリ(ライム病)
c.ボツリヌス菌
d.ペスト菌
25.1961年にメチシリン耐性黄色ブドウ球菌の最初の症例が発見されたことで、MRSAは深刻な感染症として現れてきました。MRSAを殺すことができる抗生物質が存在するのに、医療専門家はなぜそれほど心配しているのでしょうか?
a.MRSAはメチシリン耐性を他の細菌に移すことができるから。
b.患者が深刻な病気を防ぐのに十分なほど迅速に正しい抗生物質で治療されていないから。
c.MRSAが他の細菌から追加の抗生物質耐性遺伝子を獲得して「スーパーバグ」になる可能性があるから。
d.上記のすべて。
26.以下のうち、共生窒素固定を通じて起こるのはどれですか?
a.植物は無限の窒素源を使用することから利益を得る。
b.土壌は自然に施肥されることから利益を得る。
c.細菌は植物による光合成産物を使うことから利益を得る。
d.上記のすべてが起こる。
27.ある生物に見られるが、その生物内では通常は生産されていないか、または存在することが予測されていない合成化合物を_____と呼びます。
a.殺虫剤
b.バイオレメディエーター
c.難分解性化合物
d.生体異物
28.バイオレメディエーションには、_____が含まれます。
a.窒素を固定することができる原核生物の利用
b.汚染物質を浄化するための原核生物の利用
c.天然肥料としての原核生物の利用
d.上記のすべて
29.乳酸産生細菌は、ヨーグルトにその独特の風味と食感を提供することに加えて、食品生産中にどの追加の利益をもたらしますか?
a.生体異物を提供する
b.病原菌を殺すためにpHを下げる
c.乳製品を低温殺菌する
d.乳糖不耐症の人のためにラクトースを分解する
クリティカルシンキング問題
30.どのようにして環境サンプル中の培養不可能な原核生物の存在を検出するかを簡単に記述してください。
31.なぜ科学者は地球上の最初の生物が好極限性生物であったと考えているのでしょうか?
32.新しい細菌の種が発見され、岩石の中に住む好極限性生物エンドリスとして分類されました。もしこの細菌が南極大陸の永久凍土で発見された場合、この細菌が持っていなければならない2つの好極限性生物の特徴を記述してください。
33.細菌と古細菌の3つの違いに言及してください。
34.細菌と古細菌の両方の種類が同じ基本構造を持つものの、異なる化学成分から作られているという記述について説明してください。
35.ある科学者が新種の原核生物を分離しました。彼は、この標本がペプチドグリカンに対して陽性に染色する脂質二重層と細胞壁を持つ桿菌であることに気づきました。その環状染色体は単一の複製起点から複製します。この標本は古細菌、グラム陽性菌、またはグラム陰性菌のどれである可能性が最も高いですか?あなたはそれをどのようにして知ることができますか?
36.深海の熱水噴出孔で見つかるであろう条件(温度、光、圧力、有機および無機物質)について考えてみましょう。代謝ニーズ(独立栄養生物、光栄養生物、化学栄養生物など)の観点から、どのような種類の原核生物が見つかると思いますか?
37.農家は、土壌中の栄養素を維持するために、さまざまな畑で栽培されている作物を絶えず輪作しています。畑でニンジンを栽培した翌年にその畑に大豆を植えると、土壌中の窒素の維持にどのように役立ちますか?
38.土壌のある地域が汚染され、死んだ動植物を分解する細菌が死んだと想像してください。これはこの領域の炭素循環にどのように影響するでしょうか?炭素が循環のどこに蓄積するかを特定して述べてください。
39.抗生物質の無思慮で過剰な使用が世界的に大きな問題を引き起こしている理由を説明してください。
40.ホウレンソウを水で数回洗っても、大腸菌による食品媒介疾患を防ぐことができないことが研究者らによって発見されています。あなたはこの事実をどのように説明できますか?
41.あなたの友人が原核生物は常に有害で病原性があると信じています。あなたはどのようにして彼らが間違っていることを説明しますか?
42.多くの人が抗菌石鹸を使って自分の手の細菌を殺します。しかしながら、過剰使用は実際には感染のリスクを高めることがあります。これはどのように起こるのでしょうか?
解答のヒント
第22章
1 図22.8 細胞外基質と細胞の外側の層は内側の細菌を保護します。細胞が近接していることは、遺伝子の横方向伝播、すなわち抗生物質耐性遺伝子などの遺伝子をある細菌から別の細菌に導入するプロセスを促進します。そして、もし遺伝子の横方向伝播が起こらなくても、抗生物質を破壊する細胞酵素を生産する1つの細菌は近隣の細菌を救うかもしれません。3 図22.19 D 4 A 6 A 8 C 10 D 12 B 14 C 16 B 18 A 20 A 22 D 24 D 26 D 28 B 30 この生物は培養不可能であるため、その存在はPCRのような分子技術によって検出することができるでしょう。32 考えられる答えには次のものが含まれます:サイクロファイル・ハイポリス(Psychrophile Hypolith) — 低い湿度/少ない水の環境で生存するものです。34 細菌と古細菌はどちらも細胞膜を持ち、どちらも疎水性部分を含んでいます。細菌の場合、それは脂肪酸です。古細菌の場合、それは炭化水素(フィタニル)です。細菌と古細菌の両方とも、それらを保護する細胞壁を持っています。細菌の場合、それはペプチドグリカンからなりますが、古細菌の場合、それは疑似ペプチドグリカン、多糖類、糖タンパク質、または純粋なタンパク質です。細菌および古細菌の鞭毛もまた、それらの化学構造において異なります。36 回答は多岐にわたります。深海の熱水噴出孔には光がないので、原核生物は光栄養生物ではなく化学栄養生物となるでしょう。炭素源は海洋に溶け込んでいる二酸化炭素であり、そのためそれらは独立栄養生物となるでしょう。海には有機物質はそれほど多くないため、原核生物はおそらく無機物を使用することになるので、それらは化学無機栄養生物となるでしょう。熱水噴出孔では温度が非常に高いので、原核生物は好熱性でしょう。38 生態系で分解者としての役割を果たす細菌を失うと炭素循環が混乱しますが、菌類も分解者として働くことができるため、完全には停止しません。細菌の分解者が機能していなければ、有機廃棄物がその領域に蓄積し、大気中に放出される二酸化炭素が少なくなります。40 大腸菌は葉の表面にコロニーを形成し、自由な(プランクトン性の)細胞よりも除去するのが困難なバイオフィルムを形成します。さらに、細菌は植物が生育する水の中に取り込まれ、それによって単に葉の表面に存在するのではなく植物組織に侵入することができます。42 石鹸は皮膚上の細菌を無差別に殺します。これは有害な細菌を殺しますが、また皮膚から「良い」細菌を除去することもあります。非病原性細菌が除去されると、病原性細菌は空いた表面にコロニーを形成することが可能になります。
この訳文は元の本のCreative Commons BY 4.0ライセンスに従って同ライセンスにて公開します。 問題がありましたら、可能な限り早く対応いたしますので、ご連絡ください。また、誤訳・不適切な表現等ありましたらご指摘ください。この本は、https://openstax.org/details/books/biology-2e で無料でダウンロードできます。
